

Пользуясь лекциями ( на web - странице кафедры размещены презентации и текст лекций), учебниками, дополнительной литературой и другими источниками, студенты должны подготовить такие теоретические вопросы:
- Общий план строения, классификация и функциональное значение хрящевой ткани. Химический состав межклеточного вещества.
- Морфофункциональная характеристика клеточного дифферона хряща.
- Особенности строения, локализация и функции гиалинового, эластического и волокнистого хрящей.
- Строение охрястя . Его роль в питании, росте и регенерации хрящевой ткани.
- Хондрогистогенез. Интерстициальный и аппозиционный рост хрящевой ткани.
- Общий план строения, химический состав и функции костной ткани.
- Морфофункциональная характеристика клеток костной ткани.
- Особенности строения грубоволокнистой и пластинчатой костной ткани.
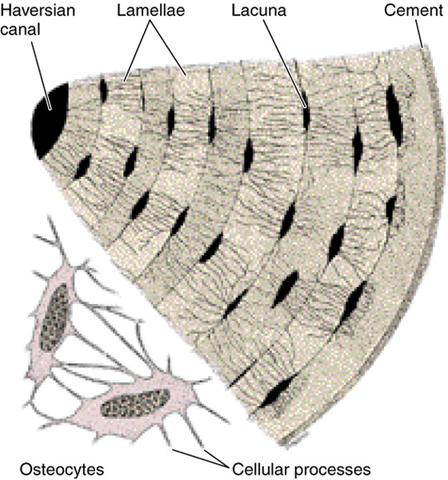
- Гистологическое строение диафиза трубчатой кости. Остеон как структурная единица кости.
- Виды остеогенеза, их характеристика.
- Рост и регенерация костей. Роль пьезоэлектрического эффекта в процессах остеосинтеза.
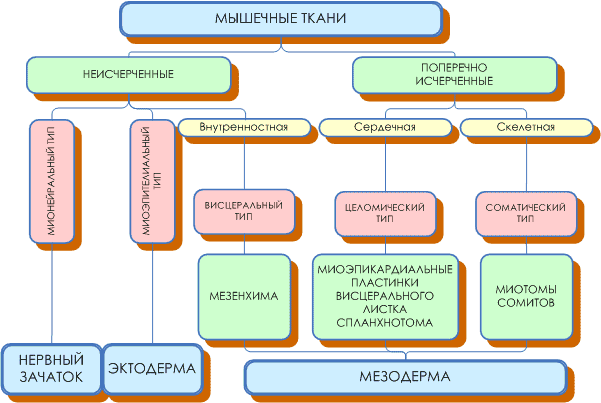
- Общая морфофункциональная характеристика мышечных тканей, источники их развития.
- Морфологическая и генетическая классификация мышечных тканей.
- Строение, локализация и особенности сокращение гладкой мышечной ткани.
- Скелетная мышечная ткань. Локализация, гистогенез, функциональные особенности.
- Гистологическое и субмикроскопическое строение волокна скелетной мышечной ткани. Саркомер.
- Особенности строения и функции сердечной мышечной ткани.
- Мышца как орган. Мион.
- Возрастные изменения и регенерация мышечных тканей.
Хрящевая ткань — разновидность соединительной ткани, состоящая из хрящевых клеток (хондроцитов) и большого количества плотного межклеточного вещества. Хондроциты имеют разнообразную форму и лежат одиночно или группами внутри хрящевых полостей. Межклеточное вещество содержит хондриновые волокна, близкие по составу к коллагеновым волокнам, и основное вещество, богатое хондромукоидом. Хрящевая ткань отличается плотным, упругим межклеточным веществом. Важнейшее отличие хрящевой ткани от костной (и большинства других типов тканей) — отсутствие внутри хряща нервов и кровеносных сосудов. Если межклеточное вещество однородно, то хрящ называется стекловидным, или гиалиновым, если пронизано волокнами — волокнистым, если заключает сеть эластических волокон — сетчатым или эластическим. Снаружи хрящ одет особой соединительнотканной оболочкой — перихондрием, или надхрястницей. Хрящ играет роль твёрдой основы, скелета тела животного или образует упругие части костного скелета (одевает концы костей, образуя суставные поверхности, или соединяет кости в виде прослоек — например, такую роль играют межпозвоночные диски.
Строительным материалом для хрящей служат хондроитин сульфат и глюкозамин. Они восстанавливают структуру суставов, эластичность суставных поверхностей и их объём, устраняют боли, связанные с гипермобильностью суставов и артрозом, усиливают действие противовоспалительных препаратов.
В основу классификации положено строение межклеточного вещества - соотношение коллагеновых и эластических волокон.
ХРЯЩЕВЫЕ ТКАНИ
гиалиновая, эластическая, волокнистая
КОСТНЫЕ ТКАНИ
грубоволокнистая, пластинчатая
Функции скелетных тканей:
- опорная;
- защитная;
- участие в вводно-солевом обмене;
- участие в минеральном обмене, депо солей кальция, фосфора и тому подобное;
- пластическая.
ХРЯЩЕВЫЕ ТКАНИ
Состав: хондробласты, хондроциты 1, 2, 3 порядка, межклеточное вещество (хондромукоид, хондриновые волокна)
http://uk.wikipedia.org/wiki/%D0%A1%D0%B2%D0%B8%D0%BD%D0%B5%D1%86%D1%8CХРЯЩЕВАЯ ТКАНЬ
Зависимо от строения межклеточного вещества различают три вида хрящевой ткани — гиалиновую, эластическую и волокнистую. Основная функция всех видов хряща — опорная, формообразовательная.
Клетки хряща - хондроциты - различных его зон характеризуются специфическими особенностями формы, положения и высоты дифференцировки. Так, непосредственно под надхрящницей локализованы незрелые хрящевые клетки - хондробласты. Они овальной формы и ориентированы длинной осью параллельно поверхности хряща. Цитоплазма этих клеток богата рибонуклеиновой кислотой, что определяет ее базофилию. В более глубоких зонах хряща хондроциты округляются или имеют неправильную многоугольную форму, их объем увеличивается.
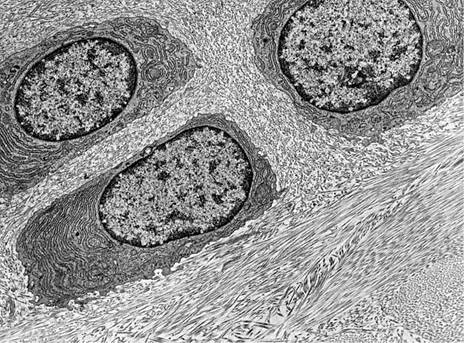
Клетки хрящевой ткани
Различают три вида хондроцитов. Первая разновидность (хондроциты І порядка) относительно малодифференцированные клетки которые находятся преимущественно в составе «молодой», так называемой первичной хрящевой ткани. Для этих клеток присущее высокое ядерно-цитоплазматическое соотношение, в цитоплазме хорошо выражены элементы комплекса Гольджи, много митохондрий и свободных рибосом. В хондроцитах второго типа ядерно-цитоплазматическое соотношения ниже, в цитоплазме повышено содержание РНК, элементов гранулярной эндоплазматической сети и комплекса Гольджи, которые обеспечивают образование и выделение в межклеточное пространство протеогликанов и гликозаминогликанов (гиалуроновой кислоты, хондроитинсульфата, кератансульфата, дерматансульфата и др). Для хондроцитов третьего типа характерный самый низкий показатель ядерно-цитоплазматического соотношения, значительное развитие гранулярной эндоплазматической сети. Интенсивность синтеза протеогликанов и гликозаминогликанов снижена сравнительно с хондроцитами второго типа.
Органические компоненты основного межклеточного вещества хрящевой ткани (хондромукоид) представлен белками, липидами, гликозаминогликанами и протеогликанами. Последние являются самой характерной особенностью хрящевой ткани. В составе протеогликанов хряща найдены гигантские макромолекулярные комплексы с молекулярной массой порядка десятков и сотен миллионов дальтон и длинной молекулы, которая составляет несколько микрометров. Они построены из длинной нити гиалуроновой кислоты, к которой нековалентными связями присоединено около сотни полипептидных цепей; со срединными аминокислотными остатками последних связано большое количество полисахаридных цепей сульфатирующих гликозаминогликанов (хондроитинсульфата, кератансульфата, дерматансульфата), а также молекул олигосахаридов. В целом молекула протеогликана напоминает веточку ели, причем от степени ее гидратации зависит упругость (тургор) хряща.
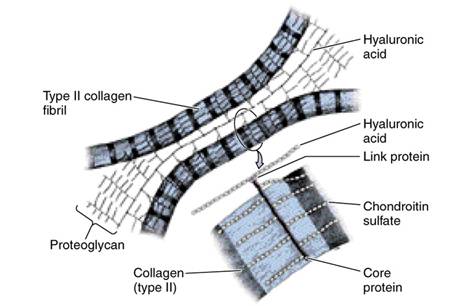
Хондриновые волокна построены из коллагена II типа (гиалиновый и волокнистый хрящи) или эластина (эластический хрящ). Ориентация волокон определяется влиянием силовых линий, которые возникают при деформации хряща в процессе функционирования органа. Особенно много хондриновых волокон вокруг лакун, которые образуют так называемую капсулу хрящевой клетки (или клеток).
Обмен веществ хряща обеспечивается циркуляцией тканевой жидкости межклеточного вещества, которая составляет до 75% общей массы ткани. Макромолекулы гликозамингликанов и протеогликанов, удерживая большое количество воды, определяют его механические свойства.
http://uk.wikipedia.org/wiki/%D0%A1%D0%B2%D0%B8%D0%BD%D0%B5%D1%86%D1%8CГиалиновый хрящ
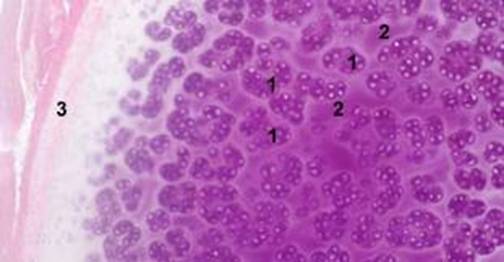
1. Изогенные групы хондроцитов; 2. Лакуны; 3. Надхрястница
Гиалиновый хрящ (textus сагrilagineus hyalinus). Локализирован в стенках трахеи, бронхов, в местах соединения ребер с грудиной, на суставных поверхностях и в метаэпифизарных пластинках роста костей. В эмбриональном периоде гиалиновый хрящ лежит в основе подавляющего большинства зачатков костей скелета. С возрастом происходит замена его костной тканью. В нативном состоянии гиалиновый хрящ бело-голубого цвета, прозрачный. Гистологически в его составе различают надхрястницу и собственно хрящ. Надхрястница состоит из поверхностного волокнистого слоя (являет собой коллагеновые волокна) и глубокого клеточного слоя (в нем содержатся хондробласты и прехондробласты). Поверхностный слой надхрястницы имеет много сосудов, которые обеспечивают трофику хряща. За счет глубокого клеточного слоя надхрястницы происходит физиологическая регенерация и апозиционный (периферийный) рост хряща. Собственно хрящ состоит из изогенных групп хондроцитов, а также молодых одиночных хондроцитов, окруженных хондромукоидом и хондриновыми волокнами. Хондромукоид, размещенный вокруг молодых хондроцитов, окрашивается оксифильно, тот который окружает более дифференцированные изогенные группы клеток, приобретает свойства базофилии. Хондриновые волокна гиалинового хряща построены из коллагена ІІ типа.
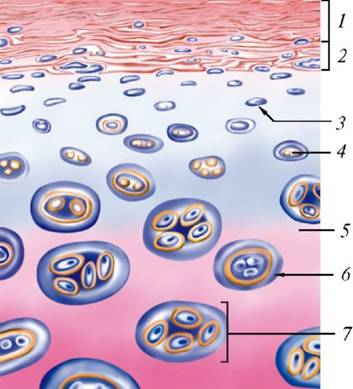
1. Поверхностный волокнистый слой надхрясницы; 2. Глубокий клеточный слой надхрясницы; 3. Хондробласты; 4. Молодые одиночные хондроциты; 5. Межклеточное вещество; 6. Хондромукоид 7. Изогенные группы клеток.

1. Поверхностный волокнистый слой надхрясницы; 2. Глубокий клеточный слой надхрясницы; 3. Хондробласты; 4. Молодые одиночные хондроциты; 5. Межклеточное вещество; 6. Изогенные группы клеток.
Эластический хрящ

1. Изогенные группы клеток; 2. Межклеточное вещество.
Эластический хрящ (textus cartilagineus elasticus) содержится в ушной раковине, слуховой трубе, внешнем слуховом проходе, клиновидных хрящах гортани. Его характерной особенностью является желтый цвет, способность растягиваться. Эластический хрящ никогда не кальцинируется. В отличии от гиалинового хряща, хондриновые волокна в эластическом хряще построены не из коллагена, а из эластина. Эластические волокна формируют капсулы вокруг хондроцитов, а также вплетаются в состав надхрястницы.

Волокнистый хрящ

Волокнистый хрящ (textus cartilagineus fibrosus) формирует межпозвонковые диски и расположен в местах перехода сухожилия в гиалиновую хрящевую ткань. Хондроциты в волокнистом хряще размещены в виде своеобразных рядов — клеточных столбиков, а коллагеновые волокна формируют толстые пластины, которые идут параллельно. По строению волокнистый хрящ напоминает сухожилие, но его клетки типично хрящевые.
Гистогенез, регенерация и вековые изменения хрящевой ткани.
В процессе эмбрионального развития зародыша мезенхима, интенсивно развиваясь, образует островки плотно прилегающих друг к другу клеток протохондральной (предхрящевой) ткани. Ее клетки характеризуются высокими значениями ядерно-цитоплазматических отношений, мелкими, плотными митохондриями, обилием свободных рибосом и слабым развитием гранулярной эндоплазматической сети. Комплекс Гольджи в клетках протохондральной ткани рассредоточен в виде небольших цистерн и пузырьков. По мере дифференцировки хондробластов они включаются в процессы синтеза макромолекулярных соединений межклеточного вещества развивающегося хряща, соответственно изменяется их синтетический и секреторный аппарат. Увеличивается объем цитоплазмы и соответственно уменьшается показатель ядерно-цитоплазматических отношений. Возрастает количество цистерн гранулярной андоплазматической сети. Комплекс Гольджи сосредоточивается вокруг ядра и расширяются его размеры. Увеличивается объем митохондрий преимущественно за счет возрастания массы их матрикса. Наблюдается выведение в окружающее межклеточное вещество содержимого вакуолей клеток. В межклеточное вещество секретируют тропоколлаген и неколлагеновые белки, а затем гликозаминогликаны и протеогликаны. Формируется первичная хрящевая (прехондральная) ткань.
Дифференцированный хондробласт морфологически характеризуется хорошо развитой гранулярной эндонлазматической сетью и обширным комплексом Гольджи. В цитоплазме клеток много включений гликогена. Увеличение массы хрящевого зачатка в процессе эмбриогенеза идет как за счет увеличения количества межклеточного вещества, так и за счет размножения хондробластов.
По мере накопления межклеточного вещества клетки развивающегося хряща изолируются в отдельных полостях (лакунах) и дифференцируются в зрелые хрящевые клетки - хондроциты.
Дальнейший рост хрящевой ткани обеспечивается продолжающимся делением хондроцитов и формированием между дочерними клетками межклеточного вещества. В более поздних стадиях развития ткани образование межклеточного вещества замедляется. Дочерние клетки, оставаясь в одной лакуне или отделяясь друг от друга лишь тонкими перегородками основного вещества, образуют характерные для зрелого хряща изогенные группы клеток (от isos - равный, одинаковый, genesis - происхождение). В дальнейшем рост хрящевой ткани обеспечивается как увеличением ее массы размножением клеток хрящевой закладки и соответственно формированием межклеточного вещества - ее интерстициальным ростом, так и продолжающимся развитием хряща за счет внутреннего - камбиального слоя надхрящницы, клетки которой, размножаясь и дифференцируясь в хондроциты, обусловливают аппозиционный рост ткани.
По мере дифференцировки хрящевой ткани интенсивность размножения клеток падает, ядра пикнотизируются, ядрышковый аппарат редуцируется.

Гистогенез хрящевой ткани
Стадии:
- Хрящевой зачаток – хондрогенный островок;
- Образование первичной хрящевой ткани ;
- Гистогенез хрящевой ткани.
Существует два способа роста хряща — внутренний (интерстиционный) и путем наложения (апозиционный). Внутренний рост хряща осуществляется в результате размножения молодых хондроцитов и новообразование изогенных групп клеток. Апозиционный рост происходит за счет надхрястницы — пролиферации хондробластов глубокого слоя, превращение хондробластов в хондроциты и продукции ими межклеточного вещества.
Физиологическая регенерация хрящевой ткани происходит благодаря деятельности хондроцитов - выработка ими веществ хондромукоида, коллагена и эластина, что способствуют новообразованию хондриновых волокон. С возрастом в хрящевой ткани уменьшается содержание клеточных элементов и растет содержание межклеточного матрикса. При этом по мере превращения хондроцитов первого и второго типов на хондроциты третьего типа, в межклеточном веществе хряща снижается количество протеогликанов, хондромукоид замещаєтся альбумоидом, растет содержание коллагеновых волокон. Последние имеют способность накапливать соли кальция. Все эти изменения приводят к уменьшению степени гидратации, потери упругости хрящевой ткани, увеличения ее ломкости. Наблюдаются также врастание в обызвествленный хрящ кровеносных сосудов и замена хрящевой ткани костной.
КОСТНЫЕ ТКАНИ
Костная ткань – особый вид соединительной ткани. Необходимо различать понятия «кость как орган» и «костная ткань»
Кость как орган – это сложное структурное образование, в которое наряду со специфической костной тканью входят надкостница, костный мозг, кровеносные и лимфатические сосуды, нервы и в ряде случаев хрящевая ткань.
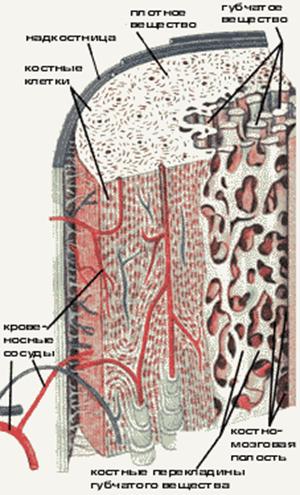
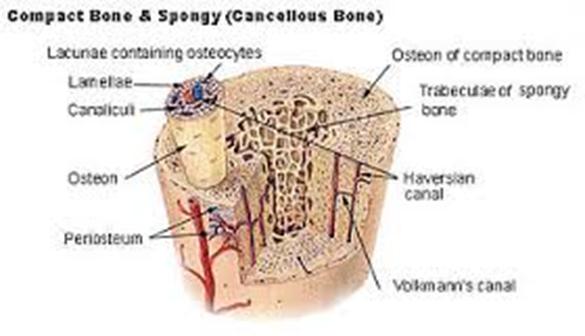
Костная ткань является главной составной частью кости. Она образует костные пластинки. В зависимости от плотности и расположения пластинок различают компактное и губчатое костное вещество. В телах длинных (трубчатых) костей в основном содержится компактное костное вещество. В эпифизах длинных костей, а также в коротких и широких костях преобладает губчатое костное вещество.

Клеточными элементами костной ткани являются остеобласты, остео-циты и остеокласты.
В состав кости входят неорганические вещества; чем младше животное, тем большее их содержание поэтому, кости молодых животных упругие и мягкие, а кости старых твердые и хрупкие. Отношение между обоими составляющими разное у всех групп позвоночных; например, в костях особенно глубоководных, содержание минеральных веществ относительно малое, и они отличаются мягким волокнистым строением.
Костная ткань (lextus osseus) вместе с хрящевой принадлежит к скелетным тканям организма. Основная роль костной ткани опорно-механическая: благодаря значительной прочности кости обеспечивают защиту жизненно важных органов от механических повреждений, а также перемещение тела в пространстве. Элементы костной ткани образуют каркас и микроокружение для клеток крови в составе красного костного мозга. Костная ткань является депо кальция и фосфора в организме. Сопротивление свежей кости на разрыв такой же меди, и в 9 раз больший, чем в свинца. Кость выдерживает сжатие 10 кг/мм2. Чтобы переломить ребро необходима сила в 110 кг/см2.
В костной ткани различают клеточные элементы (остеобласты, остеоциты и остеокласты) и межклеточное вещество (осеиновые волокна и осеомукоид). Осеомукоид содержит гликопротеины (среди которых специфический белок костной ткани остеонектин) та протеогликаны. Не обызвествленный межклеточный матрикс костной ткани имеет название остеоида (предкости).
Характерной особенностью костной ткани является исключительно высокое (до 70 %) содержание в составе ее межклеточного вещества неорганических соединений, среди которых больше всего солей кальция гидроксиапатитов и фосфатов. Прочность костей зависит от высокого содержания построенных из коллагена ІІ типа осеиновых волокон, которые образуют пластинки.
Остеобласты
Остеобласт – клетка костной ткани, участвующая в образовании межклеточного вещества. Отличительной чертой остеобластов является наличие сильно развитого эндоплазматического ретикулума и мощного аппарата белкового синтеза. В остеобластах синтезируется проколлаген, который затем перемещается из эндоплазматического ретикулума в комплекс Гольджи, включается в секретируемые гранулы (везикулы). В результате действия группы специальных пептидаз от проколлагена отщепляются сначала N-концевой, а затем С-концевой домены и формируется тропо-коллаген. Последний в межклеточном пространстве образует фибриллы. В дальнейшем после образования поперечных сшивок формируется зрелый коллаген.
В остеобластах синтезируются также гликозаминогликаны, белковые компоненты протеогликанов, ферменты и другие соединения, многие из которых затем быстро переходят в межклеточное вещество.
Остеоциты (костная клетка) – зрелая отростчатая клетка костной ткани, вырабатывающая компоненты межклеточного вещества и обычно замурованная в нем.Как известно, остеоциты образуются из остеобластов при формировании костной ткани.
Остеокласты — большие многоядерные клетки неправильной округлой формы, предшественниками которых могут быть малодифференцирированные клетки костного мозга, а также моноциты крови. Основная функция остеокластов — резорбция (рассасывание) костной ткани. Диаметр этих клеток 90 мкм и больше, в цитоплазме насчитывается от трех до нескольких десятков ядер. Цитоплазма остеокластов оксифильная или слабо базофильная, содержит значительное количество лизосом и митохондрий. На поверхности клетки, которая прикреплена к месту разрушения кости, различают две зоны: покрытую микроворсинками, зону адсорбции и секреции ферментов (так называемое гофрированное обрамление), и запирательную зону, которая изолирует участок контакта от окружающей ткани.
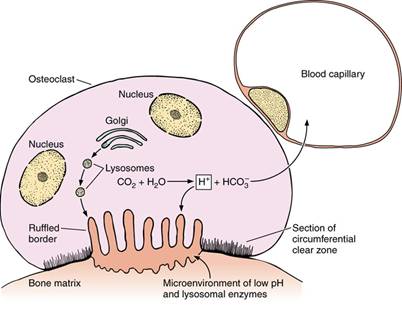
Механизм разрушительного действия остеокластов на костную ткань связывают с выделением этими клетками углекислого газа, из которого под воздействием фермента карбоангидразы образуется угольная кислота, способная растворять соли кальция. Остеоид препятствует взаимодействию остеокластов с неорганическими компонентами кости. Для реализации процесса резорбции необходима секреция остеобластами колагеназы — фермента, который разрушает слой остеоида и обеспечивает доступ остеокластов к минеральному матриксу кости.
В зависимости от способа организации коллагеновых волокон в костной ткани различают два ее вида — пластинчатую и грубоволокнистую. Для пластинчатой костной ткани характерным является строго параллельное расположение пучков коллагеновых волокон с формированием так называемых костных пластинок. В зависимости от ориентации последних в пространстве костную ткань разделяют на компактную (в ней отсутствуют полости) и губчатую (костные пластинки образуют размещенные под углом одна к одной трабекулы с формированием характерной губчатой структуры). Из компактной костной ткани построены диафизы трубчатых костей, из губчатой — плоские кости, эпифизы трубчатых костей. Пластинчатая костная ткань составляет подавляющее большинство костей организма.
Для грубоволокнистой костной ткани характерно неправильное (разнонаправленное) размещение пучков осеиновых волокон, окруженных обызвествленным осеомукоидом. Между пластинками осеиновых волокон в лакунах осеомукоида залегают остеоциты. Грубоволокнистая костная ткань встречается по большей части в скелете зародыша, а во взрослом организме — лишь в участке швов черепа и в местах прикрепления сухожилия к костям.
Строение трубчатых костей.
Диафиз это центральная часть, эпифиз — периферийное окончание трубчатых костей. В участке диафиза кости существует три слоя: надкостница (периост), собственно кость и эндост (внутренний слой). Надкостница состоит из поверхностного волокнистого слоя, образованного щепотками коллагеновых волокон, и глубокого остеогенного слоя (в нем размещены остеобласты и остеокласты). За счет надкостницы, в которой находится большое количество сосудов, осуществляется питание костной ткани; костные элементы глубокого остеогенного слоя обеспечивают рост кости в толщину, ее физиологическую и репаративную регенерацию.
Между надкостницей и собственно остеонным слоем размещен слой внешних генеральных пластинок. Основная часть стенки кости — это остеонный слой. Каждый остеон являет собой костную трубку диаметром от 20 до 300 мкм, в центральном канале которой лежит так называемый питательный сосуд и локализированы остеобласты и остеокласты. Вокруг центрального канала концентрически размещено 5-20 костных пластинок.
Между костными пластинками в костных лакунах расположены тела остеоцитов, которые анастомозируют своими отростками, размещенными в костных канальцах. Остеонный слой можно представить себе как систему параллельных цилиндров (остеонов), промежутки между которыми заполнены вставными костными пластинками. Через надкостницу к остеонному слою проходят так называемые прорывистые сосуды, а также пластинки коллагеновых волокон. От эндоста остеонный слой отделен слоем внутренних генеральных пластинок. Эндост — тонковолокнистая соединительная ткань, обогащенная остеобластами и остеокластами, которая ограничивает костномозговую полость.
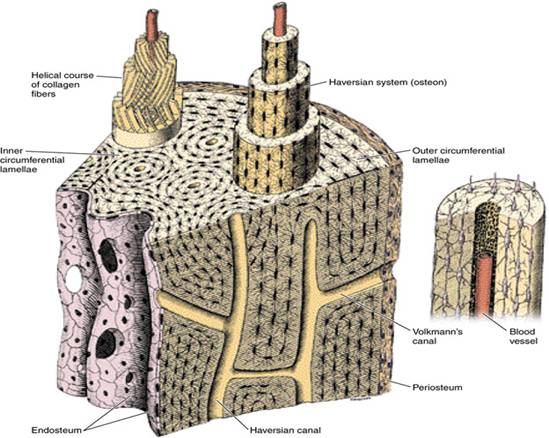
Эпифиз кости образован губчатой костной тканью. Поверхностно покрытый надкостницей, под которой размещен слой генеральных пластинок и остеонов. Костные пластинки в толще эпифиза формируют систему размещенных под углом одна к одной трабекул, полости между которыми заполнены ретикулярной тканью и гемопоетическими клетками. Подобное строение эпифиза имеют и плоские кости скелета.
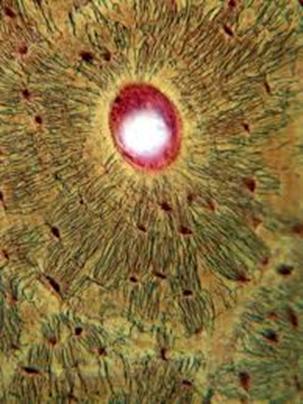
Остеон
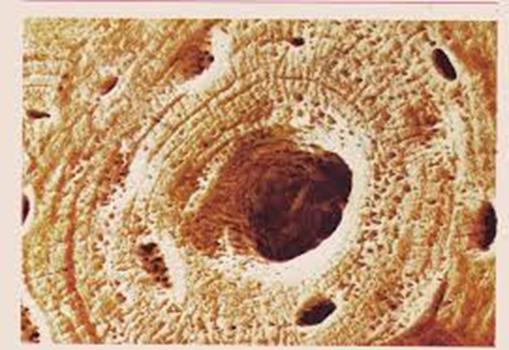
Остеон

Остеон
Одновременно с дифференцированием клеток в остеоциты образуется межклеточное вещество и коллагеновые волокна. Расположенное между клетками и волокнами основное вещество уплотняется, формируются костные балки (перекладины). Клетки поверхности образуемой кости превращаются в остеобласты. Большинство костей скелета человека (за исключением покровных костей черепа) построены из пластинчатой костной ткани, то есть имеют костные пластинки толщиной от 4 до 15 мкм, которые состоят из остеоцитов и тонковолокнистого межклеточного вещества. Соединительнотканные волокна в толще каждой пластинки лежат параллельно и ориентированы в определенном направлении. В зависимости от расположения костных пластинок различают компактные и губчатые кости. В компактной кости пластинки располагаются в определенном порядке, образовывая сложные системы остеоны.
http://uk.wikipedia.org/wiki/%D0%9C%D1%96%D0%BD%D0%B5%D1%80%D0%B0%D0%BB%D0%B8Гистогенез костной ткани.
Существует два способа развития костной ткани — непосредственно из мезенхимы (перепончатый остеогенез) и на месте хрящевого зачатка (хрящевой остеогенез). Первый способ характерен для первых недель эмбриогенеза, второй — для более поздних этапов эмбрионального развития и постнатального онтогенеза.
Из соединительной ткани развиваются кости свода и боковых отделов черепа, нижняя челюсть и, по мнению некоторых, ключица (а у низших позвоночных и некоторые другие) — это так называемые покровные или облегающие кости. Они развиваются прямо из соединительной ткани; волокна её несколько сгущаются, между ними появляются костные клетки и в промежутках между последними отлагаются известковые соли; образуются сначала островки костной ткани, которые затем сливаются между собой. Большинство костей скелета развивается из хрящевой основы, имеющей такую же форму, как будущая кость. Хрящевая ткань подвергается процессу разрушения, всасывания и вместо неё образуется, при деятельном участии особого слоя образовательных клеток (остеобластов), костная ткань; процесс этот может идти как с поверхности хряща, от одевающей его оболочки, перихондрия, превращающегося затем в надкостницу, так и внутри его. Обыкновенно развитие костной ткани начинается в нескольких точках, в трубчатых костях отдельными точками окостенения обладают эпифизы и диафиз.
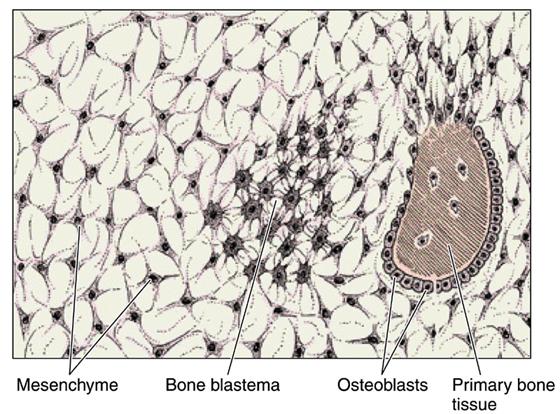
При характеристике процессов развития костной ткани непосредственно из мезенхимы определяют следующие этапы. Первый этап — формирование в составе мезенхимы так называемого остеогенного зачатка. При этом проходит локальное размножение мезенхимных клеток с врастанием в скелетогенный островок кровеносных сосудов. Остеоидный, то есть второй этап, характеризуется выделением остеогенными клетками в межклеточное пространство коллагена (формированием осеиновых волокон) и высокомолекулярных биополимеров (гликопротеинов, протеoгликанов, липидов) осеомукоида. Третий этап — собственно образование грубоволокнистой кости — заключается в известковании межклеточного вещества (откладывании солей кальция). Для реализации этого процесса необходимо присутствие в межклеточном веществе продуцируемой остеобластами щелочной фосфатазы и белка остеонектина. Последний, связывая коллаген из гидроксиапатитом, определяет место роста кристаллов фосфата кальция и их прикрепление к органичному матриксу кости. Четвертый этап связан из резорбцией ткани заключается в постоянном изменении направления действия вектора силы на кость, в результате чего возникает, так называемый пьезоэлектрический эффект (устанавливается разница потенциалов на ввогнутой и выпуклых поверхностях костных пластинок). При чем отмечено, что концентрация остеобластов и процессы апозиционного новообразования кости связаны с отрицательными зарядами, а концентрация остеокластов и процессы резорбции — с позитивными зарядами на поверхности костной ткани.
Стадии перепончатого остеогенеза:
- формирование остеогенного островка – костной бластемы;
- остеоидная – формирование осеиновых волокон и осеомукоида;
- образование грубоволокнистой костной ткани;
- ЗАМЕНА ГРУБОВОЛОКНИСТОЙ НА ПЛАСТИНЧАТУЮ КОСТНУЮ ТКАНЬ.
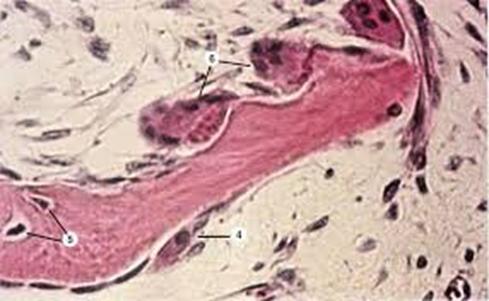
СТАДИИ ХРЯЩЕВОГО ОСТЕОГЕНЕЗА:
- образование хряща - на месте будущей кости образуется гиалиновый хрящ
- перихондральное окостенение
- проходит только в области диафиза
- в области диафиза надхрящница превращается в надкостницу, в которой появляются остеогенные клетки - остеобласты
- за счет остеогенных клеток надкостницы на поверхности хряща начинается образование кости в виде общих пластинок, имеющих циркулярный ход, наподобие годовых колец дерева (см. пластинчатую кость)
- эндохондральное окостенение
- роисходит как в области диафиза, так и в области эпифиза; окостенение эпифиза осуществляется только путем эндохондрального окостенения
- внутрь хряща врастают кровеносные сосуды, в адвентиции которых имеются остеогенные клетки - остеобласты, за счет которых вокруг сосудов происходит образование кости в виде остеонов
- одновременно с образованием кости происходит разрушение хряща
- перестройка и рост кости - старые участки кости постепенно разрушаются и на их месте образуются новые; за счет надкостницы образуются общие костные пластинки, за счет остеогенных клеток, находящихся в адвентиции сосудов кости, образуются остеоны
Метаэпифизарная пластинка имеет 4 зоны;
- неизмененного хряща:
- столбчатого хряща;
- пузырчатого хряща;
- резорбции хряща и незрелой костной ткани.
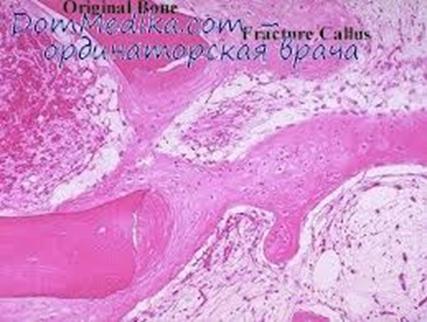
Рост кости в длину происходит главным образом в частях еще не окостенелых (в трубчатых костях между эпифизами и диафизом), но отчасти и путем отложения новых участков ткани между существующими, утолщение кости происходит путем отложения на поверхности кости новых слоев («аппозиция») благодаря деятельности остеобластов надкостницы. Последняя владеет в высокой степени способностью воспроизводить разрушенные и изъятые части кости. Деятельностью ее обусловливается и срастание переломов. Параллельно с ростом кисти идет разрушение, всасывание («резорбция») некоторых участков костной ткани, причем деятельную роль играют так называемые остеокласты («клетки, которые разрушают кисть»), многоядерные элементы, которые наблюдаются на стенках мозговых полостей, в надкостнице и стенках больших полостей в кости (напр. Гайморова пазуха и тому подобное
Изучение химического состава костной ткани сопряжено со значительными трудностями, поскольку для выделения органического матрикса требуется провести деминерализацию кости. Кроме того, содержание и состав органического матрикса подвержены значительным изменениям в зависимости от степени минерализации костной ткани.
Известно, что при продолжительной обработке кости в разведенных растворах кислот ее минеральные компоненты растворяются и остается гибкий мягкий органический остаток (органический матрикс), сохраняющий форму интактной кости. Межклеточный органический матрикс компактной кости составляет около 20%, неорганические вещества – 70% и вода – 10%. В губчатой кости преобладают органические компоненты, которые составляют более 50%, на долю неорганических соединений приходится 33–40%. Количество воды сохраняется в в тех же пределах, что и в компактной кости.
По данным А. Уайта и соавт., неорганические компоненты составляют около 1/4 объема кости; остальную часть занимает органический матрикс. Вследствие различий в относительной удельной массе органических и неорганических компонентов на долю нерастворимых минералов приходится половина массы кости.
Неорганический состав костной ткани. Более 100 лет назад было высказано предположение, что кристаллы костной ткани имеют структуру апатита. В дальнейшем это в значительной мере подтвердилось. Действительно, кристаллы кости относятся к гидроксилапатитам, имеют форму пластин или палочек и следующий химический состав – Са10(РО4)6(ОН)2. Кристаллы гидроксилапатита составляют лишь часть минеральной фазы костной ткани, другая часть представлена аморфным фосфатом кальция Са3(РО4)2. Содержание аморфного фосфата кальция подвержено значительным колебаниям в зависимости от возраста. Аморфный фосфат кальция преобладает в раннем возрасте, в зрелой кости преобладающим становится кристаллический гидроксилапатит. Обычно аморфный фосфат кальция рассматривают как лабильный резерв ионов Са2+ и фосфата.
В организме взрослого человека содержится более 1 кг кальция, который почти целиком находится в костях и зубах, образуя вместе с фосфатом нерастворимый гидроксилапатит. Большая часть кальция в костях постоянно обновляется. Ежедневно кости скелета теряют и вновь восстанавливают примерно 700–800 мг кальция.
В состав минеральной фазы кости входит значительное количество ионов, которые обычно не содержатся в чистом гидроксилапатите, например ионы натрия, магния, калия, хлора и др. Высказано предположение, что в кристаллической решетке гидроксилапатита ионы Са2+ могут замещаться другими двухвалентными катионами, тогда как анионы, отличные от фосфата игидроксила, либо адсорбируются на поверхности кристаллов, либо растворяются в гидратной оболочке кристаллической решетки.
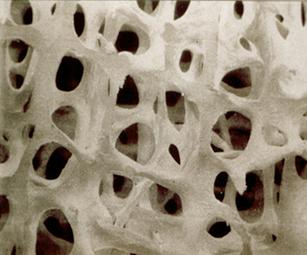
ОСТЕОПОРОЗ http://uk.wikipedia.org/wiki/%D0%9C%D1%96%D0%BD%D0%B5%D1%80%D0%B0%D0%BB%D0%B8
Вековые изменения костной ткани заключаются в постепенной потере неорганического матрикса кости после достижения двадцатилетнего возраста. Характерной особенностью является то, что у мужчин потеря минеральных компонентов кости является постоянным показателем на протяжении всей жизни и составляет около 0,4 % к массе ежегодно. У женщин с наступлением менопаузы, очевидно, в результате дефицита эстрогенов в организме, процессы деминерализации нарастают, достигая уровня 1–1,5 % ежегодно.
Количество (а точнее соотношение) органических и минеральных веществ в кости изменяется с возрастом. У детей значительно преобладают органические вещества, у взрослых минеральные. У взрослого человека часть минеральных составляющих (преимущественно, фосфорнокислая и углекислая известь, а также, хлористого кальция и другие) составляет около 60—70 % массы кости, а органичные вещества 40 %. Кости имеют большую прочность и сильное сопротивление разрыву, чрезвычайно долго противостоят разрушению. При сильном нагревании кисть теряет органические вещества, но хранит свою форму и строение; поддавая кисть действию кислоты (напр. серной), можно растворить минеральные вещества и получить гибкий хрящеватый скелет кости.
Мышечная ткань (rextus muscularis) построена из элементов, способных к сокращению, которые выполняют всю совокупность двигательных процессов в организме (крово- и лимфообращение, передвижение пищи в пищеварительном тракте, воздуха в дыхательных путях, работа сердца), а также перемещение организма или его частей в пространстве. Элементы мышечных тканей содержат специальные органеллы - миофибриллы. В их основе лежат актиновые и миозиновые миофиламенты, которые своим взаимо- действием обеспечивают процесс сокращенияи, таким образом, осуществляят функцию движения.
Существуют две классификации мышечных тканей - морфофункциональная и генетическая. Согласно морфофункциональной классификацции мышечные ткани разделяют на две группы: гладкую и поперечно - исчерчаемую, которая, в свою очередь, делится на скелетную и сердечню.
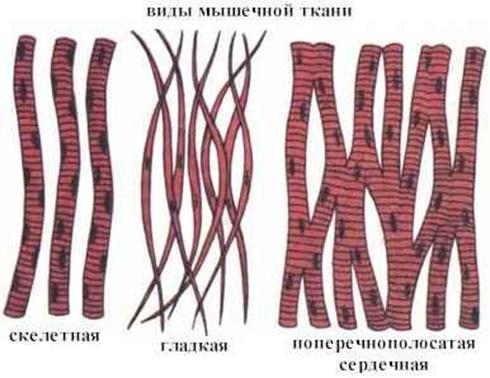
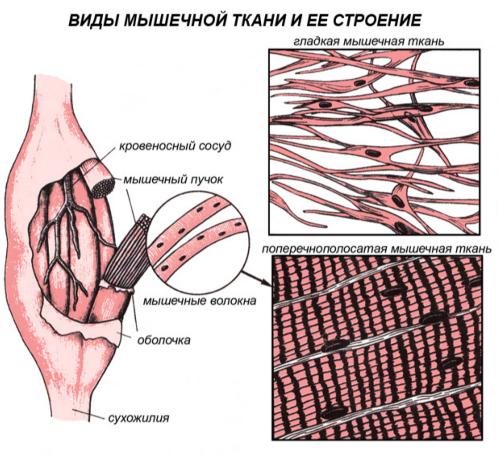
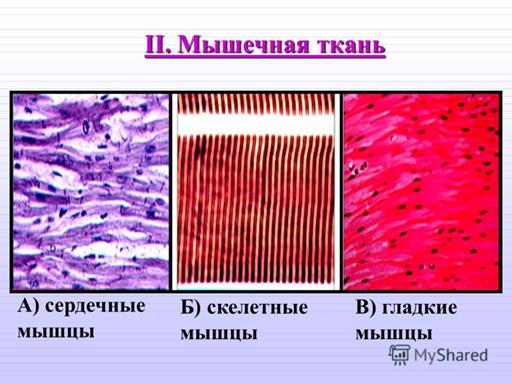
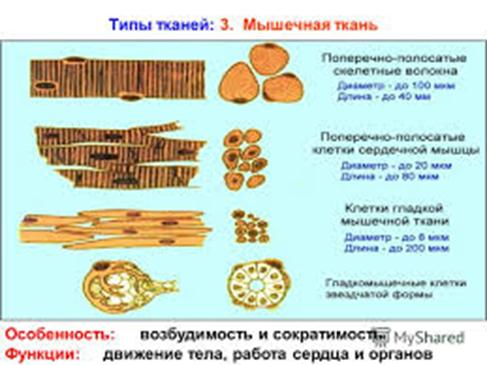
Согласно генетической классификации, которая была предложенна Н. Г. Хлопиным, мышечные ткани розделяются по их происхождению на пять гистогенетических типов:
- соматический тип происходит из миотомов мезодермы. Это скелетна мышечная ткань;
- целомический тип происходит с вентральной мезодермы. Это сердечная мышечная ткань;
- висцеральный тип происходит из мезенхимы. Это гладкая мышечная ткань внутренних органов;
- невральный тип происходит из нервной трубки. К этому типу относятся гладкие миоциты мышц радужной оболочки глаза;
- эпидермальный тип происходит из кожной эктодермы, включает миоэпителиальные корзинообразное клетки потовых, молочных, слюнных и слезных желез).
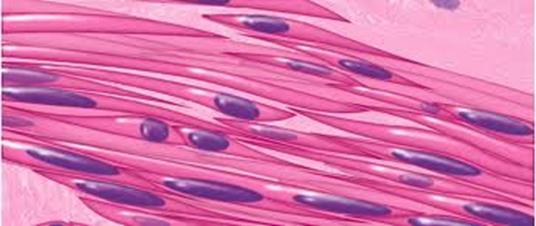
Гладкая мышечная ткань ( textus muscularis non striatus ) входит в состав стенок полостных внутренних органов, а также содержится в капсулах селезенки и лимфатических узлов, в коже. Происходит гладкая мышечная ткань из мезенхимы, то есть имеет общее происхождение с тканями внутренней среды. В группе мышечных тканей эта ткань рассматривается только с точки зрения строения и функции. Структурной единицей является гладкий миоцит.

Гладкие миоциты
Это веретенообразная клетка длинной от 20 до 100 мкм (в матке во время беременности она может достигать 500 мкм), диаметром от 2 до 20 мкм. В матке, эндокарде, аорте, мочевом пузыре встречаются миоциты с отрастками.
Ядра миоцитов паличкообразной формы и лежат в центральной широкой части клеток. Оно содержит небольшое количество гетерохроматина, хорошо заметны ядрышка. Когда миоцит сокращается, ядро изгибается. Цитоплазма окрашивается оксифильно с базофильным оттенком. Органеллы общего назначения, среди которых много митохондрий, содержатся у полюсов ядра. Комплекс Гольджи и эндоплазматическая сеть (особенно гранулярная ) развиты слабо. Цитоплазма содержит также включения - жировые, углеводные и пигментные. Цитоплазма имеет многочисленные вгинання - пиноцитозные пузырьки и кавеолы. С их помощью в цитоплазму поступают, в частности ионы кальция.
Гладкая мышечная ткань
Гладкая мышечная ткань
Миоциты не имеют поперечной исчерченности. Под электронным микроскопом в их цитоплазме выявляются тонкие актиновые миофиламенты и толстые миозиновые, расположенные преимущественно продольно, но не так как в поперечно - исчерчаемых мышцах и, очевидно, они не образовывают миофибрилл. Актиновых филаментов содержится больше. Они, кроме продольного направления, идут под углом к оси клетки, образовывая объемную сетку. Фиксируются актиновые нити к цитолеммы или друг к другу с помощью электронноплотные телец, которые состоят из белка альфа - актинина. Благодаря межмолекулярным взаимодействием с миозином актиновые нити передвигаются навстречу друг другу, тяга передается на цитолему и клетка сокращается. В механизме сокращения гладких миоцитов большую роль играет процесс фосфорилирования миозна, который зависит от концентрации ионов кальция. В свою очередь, регуляция концентрации этих ионов происходит с помощью специального белка, связывающего кальций - кальмодулина. Кальмодулин в комплексе с кальцием активирует фермент, что фосфорилирует миозин . В фосфорилированном состоянии миозин способен к взаимодействию с актином.
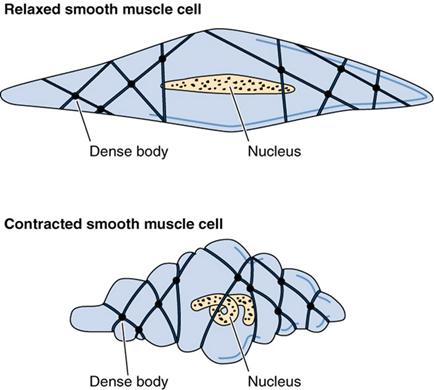
Оболочка каждого миоцита окутана тонкой базальной мембраной, к которой прикрепляются коллагеновые фибриллы. В базальной мембране имеются отверстия , в области которых мышечные клетки контактируют друг с другом с помощью щелевых контактов (нексусов). Вокруг мышечных клеток ретикулярные, эластические и тонкие коллагеновые волокна образуют сетку - эндомизий, который обьеденяет соседние миоциты. Мышечные группы по 10-12 мышечных клеток, в свою очередь, объединяются в мышечные пласты, между которыми лежит рыхлая соединительная ткань с сосудами и нервами.

Сокращается гладкая мышечная ткань ритмично, медленно, но способна долго находиться в состояние сокращения, не уставая при этом. Медленное сокращение ее обусловлено медленным циклом взаимодействия миозина с акты ном. Гладкая мускулатура способна на большие силы сокращений (например, мышечная оболочка беременной матки при родах). Тип сокращение, свойственный гладким мышцам, называется тонический. Сокращение висцеральной мускулатуры является непроизвольным, т.е. не поддается контролю сознания.
Поперечно – исчерчаемая мышечная ткань.
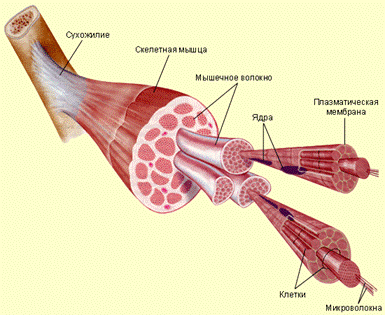
Скелетная мышечная ткань занимает 42% массы тела взрослого человека, причем подавляющее большинство мышц образована скелетной мышечной тканью. Источником развития этой ткани являются клетки миотомов дорзальной мезодермы. Эти клетки диференцирирюются в двух направлениях. Одни способны сливатся и строить симпластические структуры - мышечные трубочки, которые далее формирую дефинитивное образования - миосимпласты. Вторая линия дает при развитии - миосателитоциты.
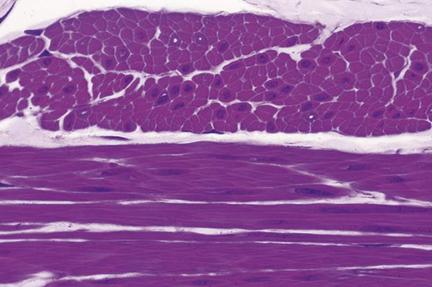
Единицей строения скелетной мышечной ткани является мышечное волокно, образованное миосимпластом и миосателитоцитамы. Мышечное волокно имеет форму цилиндра, концы которого могут быть округлены, скошенные или зазубренные. Диаметр волокна 9-150 мкм (9 мкм новорожденного ребенка, 40-50 мкм у взрослого, 150 мкм спортсмена). Длина мышечного волокна часто совпадает с длиной мышцы и может быть различна в зависимости от размеров мышцы. Например, в портняжной мышце человека она может достигать 12-13 см. Волокно окружено сарколеммой
Мышечные волокна

Мышечные волокна
Сарколемма состоит из внешней базальной мембраны. Внутренним слоем сарколеммы является плазмолемма миосимпласта. Она участвует в проведении импульсов. Между базальной мембраной и плазмолеммой симпласта расположены миосателитоциты. Это одноядерные клетки, ядра которых подобные ядер симпласта, но мелкие, круглые и ясные. Клетки имеют органеллы общего призначения, специальные органеллы отсутствуют. Миосателитоциты - это камбиальные элементы волокна, за счет которых происходит процесс роста и регенерации.
Цитоплазма симпласта имеет специальное название - саркоплазма. Ядра, численность которых может достигать нескольких десятка тысяч, как правило, лежат непосредственно под плазмолеммой, имеют удлиненно - овальную фор му, небольшое количество гетерохроматина, в них хорошо заметны ядрышки. В саркоплазме содержатся три группы организованных структуртур: общие органеллы, включенния (жировые, углеводные) и специальные органеллы - миофибриллы. Общие органеллы располагаются, главным образом, у полюсов ядер. Митохондрии крупные, многочисленные, расположены еще и между миофибриллами. Гранулярная эндоплазматическая сеть развита слабо. Агранулярная эндоплазматическая сеть развита очень хорошо, имеет специальное название саркоплазматическая сетка или саркоплазматический ретикулум, особое строение и функцию.
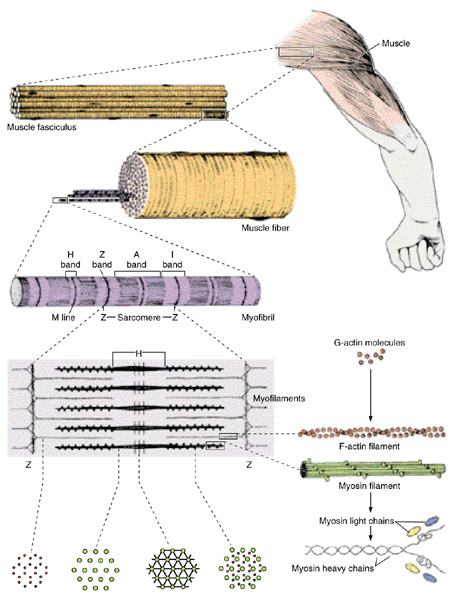
Строение миофибрилл.
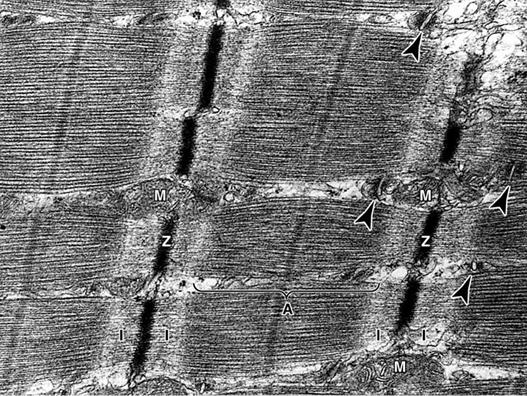
Миофибрилы расположенные вдоль мышечного волокна. Длина их совпадает с длиной мышечного волокна, толщина составляет 1-2 мкм. Миофибриллы имеют характерную поперечную исчерченность (чередование светлых и темных полос), что обусловлено особенностью их структуры и в связи с этим различными оптическими свойствами. Вследствие того, что светлые и темные полосы всех миофибрилл отдельного мышечного волокна расположены на одном уровне, все волокно является поперечно - полосатым.
В миофибрилле последовательно расположенные темные анизотропные полосы (или диски А) и светлые изотропные (или диски I). Анизотропные диски окрашиваются интенсивнее, чем изотропные. В поляризованом свете темные полосы имеют двойное променезаломливание - анизотропию, в то время как светлые полосы являются однопроменезаломлеваными ( изотропными) .

Внутри каждой I - полосы является тонкая темная линия, которая имеет название телофрагма, или линии Z. В центре темной А- зоны можно наблюдать более светлый участок - Н - зону, или полоску Гензена, на середине которой находится темная линия М, или мезофрагма. Структурной единицей миофибриллы является саркомер, который представляет собой участок между двумя телофрагмами.

Телофрагма багатая гликозаминогликанами, вследствие чего миофибриллы при мацерации обладают способностью распадатся на отдельные саркомеры (от греческого «саркос» - мясо и «мерос» - часть ). Длина саркомера составляет 2-З мкм. Структурную формулу саркомера можно записать следующим образом: Т ( Z ) + 1/ 2 І +1 / 2 А + 1/ 2 Н + Н +1 / 2 Н + 1 /2 А + 1/ 2 І + T ( Z ) . Саркомер - это элементарная сократительная единица поперечно - полосатых мышц, который сокращается благодаря тому, что может уменьшать свою длину а два раза.
Под электронным микроскопом в области саркомера были идентифицированы продольные нити, миофиламенты или микрофиламенты, двух типов - тонкие и толстые . Толстые расположенные только в средней части саркомера, построены они из белка миозина.
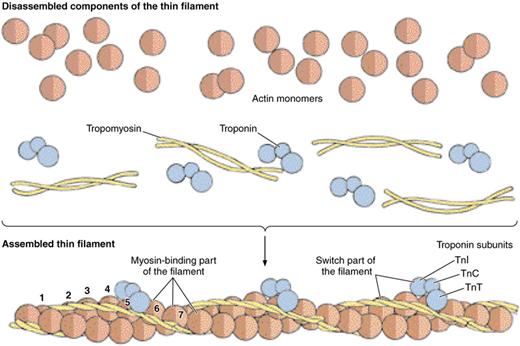
Тонкие филаменты расположены в I - полосе и частично заходят между толстыми нитями в зону Н. Одним концом они прикрепляются к телофрагме , а другой конец в них свободный, в то время как толстые филаменты имеют оба конца свободных. Тонкие филаменты построены из белка актина и, кроме того с тропомиозина и тропонина. Диаметр тонких актиновых нитей 5 нм. Толстые миозиновые нити имеют диаметр 10-12 нм и длину 1,5 мкм. Количественное отношение миозиновых нитей к актиновым 1:2 (то есть на один миозиновый миофиламент припадает два актиновых), а взаимное пространственное размещение их гексагональное: на поперечном рассечение тонкие филаменты образуют шестиугольник, в центре которого расположен толстый филамент. Если саркомер в несокращенном состоянии, наиболее темными его частями являются так называемые зоны перекрытия, то есть те части диска А, в которых есть толстые и тонкие миофиламенты. Зона Н выглядит на этом фоне светлой, так как она состоит только из толстых миозиновых нитей. При сокращении саркомера актиновые филаменты еще дальше проникают в промежуток между миозиновыми, а при полном сокращении их свободные концы почти совпадают внутрь, которые, очевидно, соединяют срединные участки соседних толстых филаментов. Электронно-микроскопические исследования показали, что Z - линия зигзагообразная, точки прикрепления тонких филаментов на одной стороне Z - пластинки лежат против промежутков между точками прикрепления таких фиментам со второй ее стороны (т.е. соседнего саркомера ). Существует мнение, что Z - пластинка построена из нитей другого типа, так называемых Z - филaмeнтов, которые сочетаются в виде решетки. Кроме того, Z - линии содержит белок (А - актинин, хотя не установлено, какие именно компоненты Z - линий построены из него.
На электронных микрофотографиях видны короткие нити, которые соединяют между собой актиновые и миозиновые филаменты, так называемые поперечные мостики. Положения их меняется при сокращении мышечного волокна.
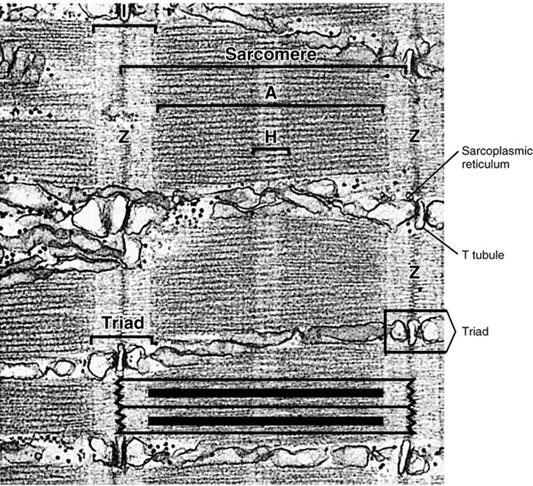
Саркоплазматическая сетка и Т -система.
Каждая манжета состоит из трех компонентов:
- терминальных цистерн (это плоские резервуары с краев манжеты);
- саркотубул (трубочек, отходящих от терминальных цистерн и идут навстречу одни к другим );
- центральной части, где саркотубулы образуют многочисленные анастомозы, напоминающие кружево. В целом описан элемент саркоплазматической сети имеет вид драной манжеты ( рваного рукава ). У млекопитающих терминальные цистерны проходят на границе А- и I- дисков саркомеров и поэтому в одном саркомере расположен один целый элемент ( манжета) на уровне диска А и половины двух соседних. Иначе говоря, элементы саркоплазматической сетки окружают А - диски, чередуются с элементами, окружающих И -диски. Элементы вокруг И - диска охватывают концевые участки соседских саркомеров.
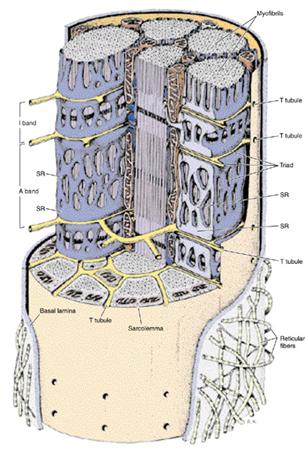
Саркоплазматическая сетка
Между двумя соседними терминальными цистернами ретикулума расположена поперечная трубочка (Т - трубочка, или Т - система). Т - трубочка - это система узких канальцев которые идут от плазмолеммы мышечного волокна (как ее вгинання) в поперечном направлении примерно на равных расстояниях. Всередине волокна T - трубочки широко разветвляются. В мышцах млекопитающих ветви двух Т - трубочек оточуют каждый саркомер на границе между А- и I- дисками и контактируют, как уже было упомянуто, с двумя терминальными цистернами саркоплазматической сетки, образуя при этом так называемую триаду. Последняя включает одну трубочку и две цистерны. Значение Т - системы заключается в том, что по ней нервный импульс плазмолеммы проникает в глубину мышечного волокна, включая все миофибриллы. Нервный импульс вызывает изменение проникливости мембран саркоплазматической сетки и выход вследствие этого ионов кальция в саркоплазму, где они необходимы для сокращения миофибрилл. Во время расслабления мышцы саркоплазматической сетки обеспечивает обратный транспорт ионов кальция от миофибрилл к своим полостям, используя для этого фермент АТФ - азу.
Молекулярные механизмы сокращения мышечного волокна.
Сучасные знания о механизме сокращения мышечного волокна базируются на представлении о филаментах двух типов, которые сдвигаются одни относительно других. Эти представления является основой модели скользящих нитей, предложенной Г. Хаксли с сотрудниками на базе электронно - микроскопических исследований. Чтобы выяснить механизм взаимодействия актиновых и миозиновых филаментов, следует рассмотреть их молекулярное строение.
Тонкий филамент являет собой двойную спираль, построенную из двух цепочек глобулярных молекул актина (остов филамента). В продольных спиральных желобках с обеих сторон от актиновых цепочек лежат молекулы тропомиозина. К молекулам тропомиозина на определенных расстояниях друг от друга присоединены молекулы тропонина. Тропомиозин вместе с тропонином играет основную роль в регуляции взаимодействия актина с миозином.
олстые филаменты состоят из молекул миозина. Каждая молекула имеет двойную головку и длинный хвост и может сгибаться в двух местах так, что головка и проксимальная часть хвоста способны вращатся, как на шарнире. В толстом филаменте молекулы миозина лежат параллельно, образуя пучок. Половина их обращена головками к одному концу филамента, а вторая - в другой. Молекулы миозина несколько сдвинуты друг относительно друга и их головки распологаются вдоль толстого филамента, исключая его серединную часть, где головок нет совсем.
Срединная часть толстого филамента построена только из хвостов миозиновых молекул. На электронных микрофотографиях головкам молекул миозина отвечают вышеупомянутые поперечные мостики, которые во время сокращения мышечного волокна образуют многочисленные соединения между толстыми и тонкими филаментами. Головки миозина расположенны по спирали, образуя шесть продольных рядов. Каждый ряд головок лежит точно против одного из шести тонких филаментов, которые окружают один толстый филамент. Во время сокращения, головки миозина присоединяются к молекулам актина в соседнем тонком филаменте.
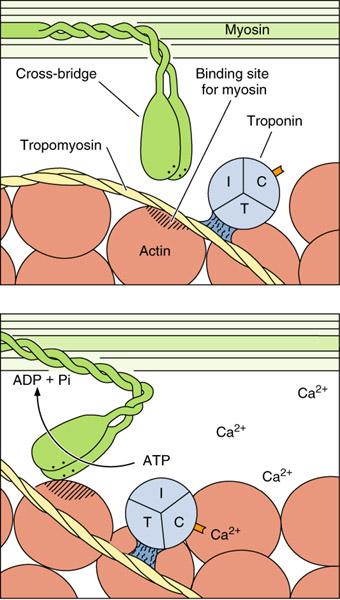
Комплексы тропонина и тропомиозина действуют как своеобразное молекулярное «замыкальное устройство», которое во время расслабления мышечного волокна не дает молекулам актина взаимодействовать с миозиновыми головками толстых филаментов. «Открывают» актин ионы каль- ция, которые освобождаются из полостей саркоплазматической сети при распространении импульса по Т - трубочках. После остановки стимуляции ионы кальция быстро транспортируются от миофибрилл к саркоплазматической сетки. Тогда актин снова замыкается и сокращения прекращаются. Механизм, с помощью которого ионы кальция «открывают» актин, связанный с их присоединением к тропонину: молекулы тропомиозина при этом сдвигаются и открывают участки актина, способные взаимодействовать с головками миозина.
Энергию, необходимую для сокращения мышц, дает АТФ. Головки миозина способны связывать молекулы АТФ и имеют АТФ - азную активность (способны расщеплять АТФ). Энергия, которая высвобождается при этом используется на изгиб молекул миозина в «шарнирных» участках, их присоединение к актиновым филаментам и продвижение последних вдоль миозиновых. Комплекс актина с миозином и АТФ не стабилен и быстро розпадается на актин и миозин - АТФ. Очевидно, поперечные мостики отделяются в тот момент, когда головки миозина связываетют молекулы АТФ. Согласно рассчетам этот цикл повторяется с огромной скоростью - 50-100 раз в секунду. Интересным есть тот факт, что после смерти, вследствие прекращения синтеза АТФ, в мышцах не остается молекул, которые вызвали б отделение миозина от актина и актиномиозиновий комплекс стабилизируется на несколько часов. Филаменты фиксируются в соединеном положении. Это состояние имеет название трупного одубивания и сохраняется к появлению аутолитических изменений, после чего мышцы становятся способными к пассивному расслаблению.
Красные и белые мышечные волокна.
В саркоплазме содержится растворимый пигментный белок миоглобин . По своему химическому строению этот белок очень близок к гемоглобину крови и также способен связывать кислород и отдавать его при необходимости. Миоглобин окрашивает мышечные волокна в красный цвет. Зависимо от содержания саркоплазмы (а, следовательно, и миоглобина), толщины и ферментного состава мышечные волокна подразделяют на красные, белые и промежуточные . Мышцы человека в основном содержат все три типа волокон, но их соотношения зависит от функции того или иного мышца. Красные волокна имеют незначительную толщину, большое количество миоглобина в саркоплазме, многочисленные митохондрии, богатые цитохромы. Белые волокна толще, они содержат меньше миоглобина и митохондрий. Волокна третьего типа занимают промежуточное положение по этим показателям. Мышцы, в которых преобладают красные волокна, способны к более длительной активности, чем мышцы, состоящие преимущественно из белых волокон. Потому что их саркоплазма хорошо приспособлена к обеспечению своих энергетических потребностей. Белые волокна способны сокращатся быстрее, чем красные, но они сравнительно быстро устают, так как не могут долго получать достаточное количество энергии.
Функциональные особенности поперечно – исчерчаемой мышечной ткани.
С поперечно - исчерчаемой мышечной ткани построены мышцы скелета человека, сокращение которых зависит от сознания, в отличие от непроизвольного сокращения гладких мышц. Поперечно - исчерчаемым мышцам присущ так называемый тетанических тип сокращения, для которого характерны такие признаки: сокращение сильные, быстрые (сокращение мышечных волокон в 10-25 раз быстрее, чем гладких мышечных клеток), не продолжительны. Исчерчаемые мязы быстрее устают и не могут находиться в состоянии сокращения так долго, как гладкие.
Строение мышцы как органа.
Отдельные поперечно - исчерчаемые мышечные волокна сочетаются соединительной тканью в орган, имеющий название мышцы. Тонкие слои рыхлой соединительной ткани между мышечными волокнами называют эндомизием. Ретикулярные и коллагеновые волокна эндомизия переплетаються с волокнами сарколеммы. На конце каждого мышечного волокна плазмолемма образует узкикие глубокие вгинання, в которые про- никают коллагеновые и ретикулярные волокна.
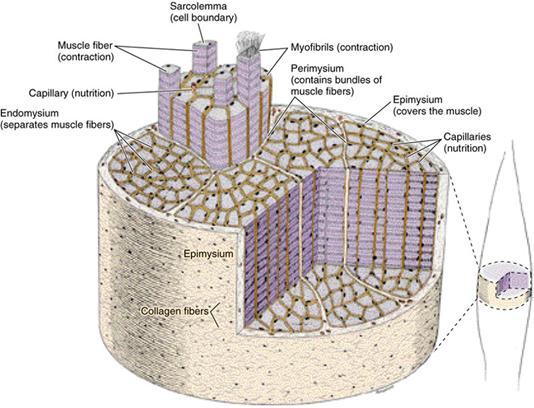
Последние пронизывают базальную мембрану и образуют петлю, которая фиксируется к плазмолемме именно в том месте, где с ней контактируют актиновые нити саркомеров. После выхода из базальной мембраны ретику лярные волокна переплетаются с коллагеновыми, а последние переходят в сухожилия. Каждое мязовое волокно имеет самостоятельную иннервацию и окружено сеткой гемокапилляров. Комплекс волокна с окружающими его элементами рыхлой соединительной ткани является структурной и функциональной единицей скелетной мышцы и называется мионом.
Сердечная мышечная ткань
Гистогенез сердечной мышечной ткани. Источники развития сердечной мышечной ткани находятся в прекардиальной мезодерме. В гистогенезе возникают парные складчатые утолщения висцерального листка спланхнотома — миоэпикардиальные пластинки, содержащие стволовые клетки сердечной мышечной ткани. Последние путем дивергентной дифференцировки дают начало следующим клеточным дифферонам: рабочим, ритмзадающим (пейсмекерным), проводящим и секреторным кардиомиоцитам. Исходные клетки сердечной мышечной ткани — кардиомиобласты характеризуются рядом признаков: клетки уплощены, содержат крупное ядро, светлую цитоплазму, бедную рибосомами и митохондриями. В дальнейшем происходит развитие комплекса Гольджи, гранулярной эндоплазматической сети. В кардиомиобластах обнаруживаются фибриллярные структуры, но миофибрилл нет. Клетки обладают высоким пролиферативным потенциалом. После ряда митотических циклов кардиомиобласты дифференцируются в кардиомиоциты, в которых начинается саркомерогенез. В цитоплазме кардиомиоцитов увеличивается число полисом, канальцев гранулярной эндоплазматической сети, накапливаются гранулы гликогена, возрастает объем актомиозинового комплекса. Кардиомиоциты сокращаются, но не теряют способность к дальнейшей пролиферации и дифференцировке. Развитие сократительного аппарата в позднем эмбриональном и постнатальном периодах происходит путем надставки новых саркомеров и наслоения вновь синтезированных миофиламентов. Дифференцировка кардиомиоцитов сопровождается увеличением числа митохондрий, распределением их у полюсов ядер и между миофибриллами и протекает параллельно со специализацией контактирующих поверхностей клеток. Кардиомиоциты путем контактов "конец в конец", "конец в бок" формируют клеточные комплексы — сердечные мышечные волокна, и в целом ткань представляет собой сетевидную структуру.
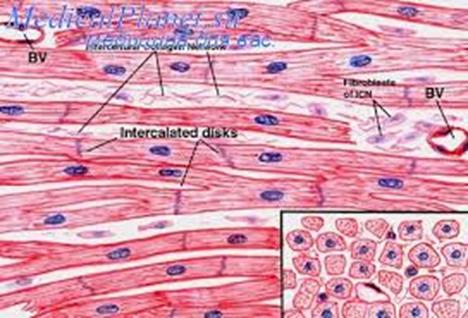
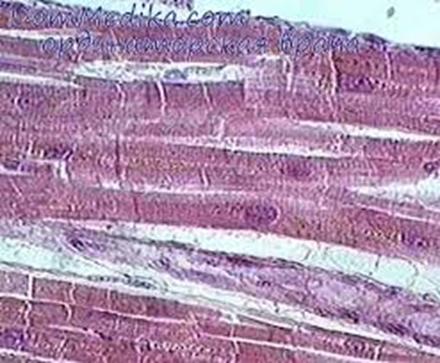
Строение сердечной мышечной ткани. Структурно-функциональной единицей волокон есть кардиомиоциты — это клетки, имеющие вытянутую прямоугольную форму. Длина рабочих кардиомиоцитов составляет 50-120 мкм, а ширина — 15-20 мкм. Одно-два ядра располагаются в центре клетки.
Сердечная мышечная ткань
Сердечная мышечная ткань
Периферическую часть цитоплазмы кардиомиоцитов занимают поперечноисчерченные миофибриллы, аналогичные таковым в симпластах скелетномышечного волокна. Однако каналы саркоплазматической сети и Т-системы менее отчетливо выражены. Кардиомиоциты отличаются большим количеством митохондрий, расположенных тесными рядами между миофибриллами. Снаружи миоциты покрыты сарколеммой, в составе которой выделяются плазмолемма и базальная мембрана. Характерной особенностью ткани является наличие вставочных дисков на границе между контактирующими кардиомиоцитами. Вставочные диски пересекают волокно в виде волнистой или ступенчатой линии и включают межклеточные контакты от простых, по типу десмо-сом и до щелевых (нексусов).
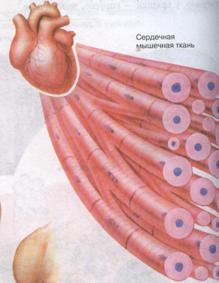
Сердечная мышечная ткань
http://uk.wikipedia.org/wiki/%D0%9C%D1%96%D0%BD%D0%B5%D1%80%D0%B0%D0%BB%D0%B8Источники информации
- Улумбеков Э.Ф., Чельшева Ю.А. Гистология, эмбриология. Цитология / Э.Ф. Улумбеков, Ю.А. Чельшева – М. : ГЕО ТАР. – Медиа, 2007. – С. 145 - 192
- Данилов Р. К. Гистология. Эмбриология. Цитология. : [учебник для студентов медицинских вузов] / Р. К. Данилов – М. : ООО «Медицинское информационное агенство», 2006. – С. 152 – 177.
- Гистология, цитология и эмбриология / [Афанасьев Ю. И., Юрина Н. А., Котовский Е. Ф. и др.] ; под ред. Ю. И. Афанасьева, Н. А. Юриной. – [5-е изд., перераб. и доп.]. – М. : Медицина. – 2001. – С. 224 - 268
- Кузнецов С. Л. Атлас по гистологии, цитологии и эмбриологии / Кузнецов С. Л., Н. Н. Мушкамбаров, В. Л. Горячкина. – М. : Медицинское информационное агенство, 2002. – С. С. 81 - 98. http://uk.wikipedia.org/wiki/%D0%97%D0%BE%D0%B1%D1%80%D0%B0%D0%B6%D0%B5%D0%BD%D0%BD%D1%8F:Bone-inside.jpg - http://uk.wikipedia.org/wiki/%D0%97%D1%83%D0%B1
{kind=link}
Автор доц. Шутурма О.Я.http://uk.wikipedia.org/wiki/%D0%97%D1%83%D0%B1
http://uk.wikipedia.org/w/index.php?title=%D0%93%D0%B0%D0%B9%D0%BC%D0%BE%D1%80%D0%BE%D0%B2%D0%B0_%D0%BF%D0%B5%D1%87%D0%B5%D1%80%D0%B0&action=edit http://uk.wikipedia.org/w/index.php?title=%D0%93%D0%B0%D0%B9%D0%BC%D0%BE%D1%80%D0%BE%D0%B2%D0%B0_%D0%BF%D0%B5%D1%87%D0%B5%D1%80%D0%B0&action=edit http://uk.wikipedia.org/w/index.php?title=%D0%93%D0%B0%D0%B9%D0%BC%D0%BE%D1%80%D0%BE%D0%B2%D0%B0_%D0%BF%D0%B5%D1%87%D0%B5%D1%80%D0%B0&action=edit