

Biochemistry of hormones: сlassification, mechanism of influence at target cells. Biochemistry of thyroid and parathyroid glands hormones. Introduction To metabolism. General pathways of metabolism in the organism. Bioenergetics. Krebs cycle, biological oxidation, oxidative phosphorylation.
The survival of multicellular organisms depends on their ability to adapt to a constantly changing environment. Intercellular communication mechanisms are necessary requirements for this adaptation. The nervous system and the endocrine system provide this intercellular, organism- wide communication. The nervous system was originally viewed as providing a fixed communication system, whereas the endocrine system supplied hormones, which are mobile messages. In fact, there is a remarkable convergence of these regulatory systems. For example, neural regulation of the endocrine system is important in the production and secretion of some hormones; many neurotransmitters resemble hormones in their synthesis, transport, and mechanism of action; and many hormones are synthesized in the nervous system.
Endocrine vs. Nervous System
|
Nervous System |
Endocrine System |
|
Neurons release neurotransmitters |
Endocrine cells release hormones |
|
A neurotransmitter acts on specific cell right next to it. |
Hormones travel to another nearby cell or act on cell in another part of the body. |
|
Neurotransmitters have their effects within milliseconds. |
Hormones take minutes or days to have their effects. |
|
The effects of neurotransmitters are short-lived. |
The effects of hormones can last hours, days, or years. |
|
Performs short term crisis management |
Regulates long term ongoing metabolic function |
|
Neurotransmitter acts on specific cell right next to it. |
Hormone can travel to another nearby cell or it can act on another part of the body. |
The word “hormone” is derived from a Greek term that means to arouse to activity. As classically defined, a hormone is a substance that is synthesized in one organ and transported by the circulatory system to act on another tissue. They are secreted in response to changes in the environment inside or outside the body. These are released into the extracellular fluid, where they are diffused into the blood stream. The latter carries them from the site of production to the site of action. They act on specific organs called target organs. The blood contains all the hormones but the cells of a target organ can pick up the specific required hormone only and ignore all others. It has been found that the target cell has on its surface or in its cytoplasm a specific protein molecule, called a receptor, which can recognise and pick out the specific hormone capable of action in that cell. The hormone delivers its message to the target cell by changing the shape of the receptor cell and binds to it. The receptors new shape sets up certain changes in the cell such as alteration in permeability, enzyme activity or gene transcription.
http://www.youtube.com/watch?v=kIPYVV4aThM&feature=related

Hormones may stimulate or inhibit specific biological processes in the target organs to modify their activities thus acting as regulators. There is considerable co-ordination between nerves and hormones. Nerves regulate synthesis and release of some hormones. Some times hormones may also influence nerve activities. Thus, hormonal co-ordination plays an important role in regulating body functions.
Calcitonin secreted by thyroid gland regulates the concentration of calcium and phosphorus in the blood. When the concentration of calcium rises in the blood, the secretion of calcitonin is seen which lowers the concentration of calcium and phosphorus in the plasma by decreasing the release for the bones.
Feedback controls
Maintenance of internal chemical environment of the body to a constant is called homeostasis. Hormones play a major role in maintaining homeostasis by their intergrated action and feed back controls.
Feedback control is mostlly negative, rarely positive. In a negative feedback control, synthesis of a hormone slows or halts when its level in the blood rises above normal. Some of examples of feedback control is given below.
Rise of testosterone level in the blood above normal inhibits ICSH secretion by the anterior pituitary lobe. This negative feedback checks oversecretion of testosterone
Hypothalamus in response to some external stimulus, produces a thyrotrophin-releasing hormone for the secretion of thyrotrophic hormone. The thyrotrophin-releasing hormone (TRH) stimulates the anterior pituitary lobe to secrete thyrotrophic hormone. The latter in turn stimulates the thyroid gland to produce thyroxine. If thyroxine is in excess, it exerts an influence on the hypothalamus and anterior pituitary lobe, which then secrete less releasing hormone and thyroid-stimulating hormone (TSH) respectively. A rise in the TSH level in the blood may also exert negative feed back effect on the hypothalmus and retard the secretion of TRH. This restores the normal blood-thyroxine level.
Functions of hormones.
Hormones regulate the following processes:
Growth and differentiation of cells, tissues, and organs These processes include cell proliferation, embryonic development, and sexual differentiation— i. e., processes that require a prolonged time period and involve proteins de novo synthesis. For this reason, mainly steroid hormones which function via transcription regulation are active in this field
Metabolic pathways Metabolic regulation requires rapidly acting mechanisms. Many of the hormones involved therefore regulate interconversion of enzymes. Themain processes subject to hormonal regulation are the uptake and degradation of storage substances (glycogen, fat), metabolic pathways for biosynthesis and degradation of central metabolites (glucose, fatty acids, etc.), and the supply of metabolic energy.
Digestive processes Digestive processes are usually regulated by locally acting peptides (paracrine), but mediators, biogenic amines, and neuropeptides are also involved.
Maintenance of ion concentrations (homeostasis) Concentrations of Na+, K+, and Cl– in body fluids, and the physiological variables dependent on these (e. g. blood pressure), are subject to strict regulation. The principal site of action of the hormones involved is the kidneys, where hormones increase or reduce the resorption of ions and recovery of water. The concentrations of Ca2+ and phosphate, which form the mineral substance of bone and teeth, are also precisely regulated. Many hormones influence the above processes only indirectly by regulating the synthesis and release of other hormones (hormonal hierarchy).
3. Endocrine, paracrine, and autocrine hormone effects.
http://www.youtube.com/watch?v=3LW7TSBcFjE&feature=related

Hormones transfer signals by migrating from heir site of synthesis to their site of action. They are usually transported in the blood. In this case, they are said to have an endocrine effect (example: insulin). By contrast, tissue hormones, the target cells for which are in the immediate vicinity of the glandular cells that produce them, are said to have a paracrine effect (example: gastrointestinal tract hormones). When signal substances also pass effects back to the cells that synthesize them, they are said to have an autocrine effect (example: prostaglandins). Autocrine effects are often found in tumor cells, which stimulate their own proliferation in this way. Insulin,which is formed in the B cells of the pancreas, has both endocrine and paracrine effects. As a hormone with endocrine effects, it regulates glucose and fat metabolism. Via a paracrinemechanism, it inhibits the synthesis and release of glucagon from the neighboring A cells.
Endocrine glands
The endocrine system is made up of glands that produce and secrete hormones, chemical substances produced in the body that regulate the activity of cells or organs. These hormones regulate the body's growth, metabolism (the physical and chemical processes of the body), and sexual development and function. The hormones are released into the bloodstream and may affect one or several organs throughout the body.
The major glands of the endocrine system are the hypothalamus, pituitary, thyroid, parathyroids, adrenals, pineal body, and the reproductive organs (ovaries and testes). The pancreas is also a part of this system; it has a role in hormone production as well as in digestion.
The endocrine system is regulated by feedback in much the same way that a thermostat regulates the temperature in a room. For the hormones that are regulated by the pituitary gland, a signal is sent from the hypothalamus to the pituitary gland in the form of a "releasing hormone," which stimulates the pituitary to secrete a "stimulating hormone" into the circulation.
The stimulating hormone then signals the target gland to secrete its hormone. As the level of this hormone rises in the circulation, the hypothalamus and the pituitary gland shut down secretion of the releasing hormone and the stimulating hormone, which in turn slows the secretion by the target gland. This system results in stable blood concentrations of the hormones that are regulated by the pituitary gland.
Classification of hormones.
The animal organism contains more than 100 hormones and hormone-like substances, which can be classified either according to their structure or according to their function. In chemical terms, most hormones are:
Ø hormones of protein structure: all hormones of anterior pituitary (except ACTH), insulin, parathyroid hormone;
Ø hormones of peptide structure: ACTH, calcitonin, glucagon, hormones of posterior pituitary, factors of hypothalamus, thymozin;
Ø steroid hormones: adrenal cortical steroids, sex hormones;
Ø
Ø hormones - derivatives of amino acid: thyroid hormones, adrenal medulla hormones, epiphysis hormones;
Ø
Ø hormones derivatives of unsaturated fatty acid: prostaglandins.
Pathways in biosynthesis of eicosanoids from arachidonic acid: there are parallel paths from
v Lipotrophic
v Hydrophilic
http://www.youtube.com/watch?v=kIPYVV4aThM&feature=related
http://www.youtube.com/watch?v=8fh2HmdxQjQ&feature=related
Mechanism of action
A. Mechanism of action of lipophilic hormones
Lipophilic signaling substances include the steroid hormones, calcitriol, the iodothyronines (T3 and T4), and retinoic acid. These hormones mainly act in the nucleus of the target cells, where they regulate gene transcription in collaboration with their receptors and with the support of additional proteins (known as coactivators and mediators). There are several effects of steroid hormones that are notmediated by transcription control. These alternative pathways for steroid effects have not yet been fully explained. In the blood, there are a number of transport proteins for lipophilic hormones. Only the free hormone is able to penetrate the membrane and enter the cell. The hormone encounters its receptor in the nucleus (and sometimes also in the cytoplasm). The receptors for lipophilic hormones are rare proteins. They occur in small numbers (103–104 molecules per cell) and show marked specificity and high affinity for the hormone (Kd = 10–8–10–10 M). After binding to the hormone, the steroid receptors are able to bind as homodimers or heterodimers to control elements in the promoters of specific genes, from where they can influence the transcription of the affected genes—i. e., they act as transcription factors. The illustration shows the particularly well-investigated mechanism of action for cortisol, which is unusual to the extent that the hormone–receptor complex already arises in the cytoplasm. The free receptor is present in the cytoplasm as a monomer in complex with the chaperone hsp90. Binding of cortisol to the complex leads to an allosteric conformational change in the receptor, which is then released from the hsp90 and becomes capable of DNA binding as a result of dimerization. In the nucleus, the hormone–receptor complex binds to nucleotide sequences known as hormone response elements (HREs).
The second mechanism involves steroid hormones, which pass through the plasma membrane and act in a two step process. Steroid hormones bind, once inside the cell, to the nuclear membrane receptors, producing an activated hormone-receptor complex. The activated hormone-receptor complex binds to DNA and activates specific genes, increasing production of proteins.
|
|
|
|
These are short palindromic DNA segments that usually promote transcription as enhancer elements. The illustration shows the HRE for glucocorticoids (GRE; “n” stands for any nucleotide). Each hormone receptor only recognizes its “own” HRE and therefore only influences the transcription of genes containing that HRE. Recognition between the receptor and HRE is based on interaction between the amino acid residues in the DNA-binding domain (B) and the relevant bases in the HRE (emphasized in color in the structure illustrated). As discussed on p. 244, the hormone receptor does not interact directly with the RNA polymerase, but rather—along with other transcription factors—with a coactivator/mediator complex that processes all of the signals and passes them on to the polymerase. In this way, hormonal effects lead within a period of minutes to hours to altered levels ofmRNAs for key proteins in cellular processes (“cellular response”).
B. Mechanism of action of hydrophilic hormones
The hormones are released in very small quantities, yet they can cause widespread dresponses in cells or tissues all over the body. These responses in cells or tissues all over the body. These responses can be quite specific and selective in different cells. All vertebrate hormones belong to one of four chemical groups. Some hormones, such hormone, such as adrenaline and thyroid hormone, are small molecules derived from the amino acid tyrosine, others such as vasopressin and oxytocin, are short peptides, still other hormones, like insulin and glucagons, are longer polypeptide chains. Testosterone and estrogen are steroid hormones. Catecholamines, peptide and protein hormones are not lipid-soluble, and so, cannot enter their target cells through the bilipid layer of plasma membrane. Instead, these water-soluble hormones interact with a surface receptor, usually a glycoprotein, and thus, initiate a chain of events within it. The hormone insulin provides a well-studied example of how this happens.
Extracellular Receptor
The membrane bound receptors of insulin is a heterotetrameric protein consisting of four subunits, two -subunits protrude out from surface of the ell and bind insulin, and two -subunits that span the membrane and protrude into the cytoplasm.
Such
receptors range from fewer than
1) Binding to the receptor
Binding of insulin to the outer subunits of the receptor causes a conformational change in the membrane spanning -subunits, which is also an enzyme, a tyrosine kinase. The activated -subunits add phosphate groups of specific tyrosine residues located in cytoplasmic domain of the receptor, as well as a variety of insulin receptor substrates.
2) Second messengers the mediator
As a result of -subunit activity, a transducer G protein activates enzyme phosphodiesterase. This enzyme makes phosphatidylinositol 4,5-biphosphate (PIP2) into a pair of mediators inositoltriphosphate (IP3) and diacylglycerol (DG). In turn, IP3, which is water-soluble, and so diffuses into cytoplasm triggers the release of another messenger Ca2+ ions from intracellular endoplasmic reticulum activating many calcium-mediated processes. While DG remains in the membrane where it activates an enzyme called protein kinase C, which in turn, activates many other enzymes, such as pyruvate dehydrogenase, and so brings about the physiological effects.
3) Amplification of signal
Mediators amplify the signal in an expanding cascade of response. A single -subunit of insulin receptor, for example, activates many molecules of DG, and each protein kinase C molecule activated by DG will, in turn, activate many other enzyme molecules. DG and IP3 are examples of second messengers, intermediary compounds that amplify a hormonal signal and so set into action a variety of events within the affected cell. A variety of events within the affected cell. A variety of hormones use another second messenger, the cyclic form of adenosine monophosphate, (cAMP). The enzyme adenylate cyclase converts adenosine triphosphate (ATP) into cAMP. Because an enzyme can be used over and over again, a single molecule of active adenylate cyclase can catalyse production of about 100 molecules of cAMP. In muscle or liver cells, when hormones, such as, adrenaline bind receptors, the receptors change shape and bind to G protein, causing it, in turn, to bind the nucleotide guanosine triphosphate (GTP) and activate another protein adenylate cyclase. The result of this complex cascade of interactions is the production of large amounts of cAMP.
cAMP activates the enzyme protein kinase A, which, in turn, activates the enzyme phosphorylate kinase. Each molecule of protein kinase A activates roughly 100 molecules of enzyme, phosphorylate kinase and so on. The net result is that a single molecule of adrenaline may lead to release of as many as 100 million molecules of glucose within only 1 or 2 minutes. No wonder only very small quantities of hormone are needed.
4) Antagonistic effect
Many cells use more than one second messenger. In heart cells, cAMP serves as a second messenger, speeding up muscle cell contraction in response to adrenaline, while cyclic guanosine monophosphate (cGMP) serves as another second messenger, slowing muscle contraction in response to acetylcholone. It is in this way that the sympathetic and parasympathetic nervous systems achieve antagonistic effect on heartbeat. Another example of antagonistic effect is insulin, which lowers blood sugar level, and glucagons, which raises it.
5) Synergistic effect
Another type of hormonal interaction is known as synergistic effect. Here, two or more hormones complement each others actions and both are needed for full expression of the hormone effects. For example, the production, secretion and ejection of milk by mammary glands require the synergistic effects of estrogens, progesterone, prolactin and oxytocin.
Intracellular Receptors
We have discussed many dramatic effects of hormone, for instance, testosterone. Yet, its concentration in the plasma of adult human male is only 30 to 100 ng per ml. How can hormones in such tiny quantities have such widespread and selective actions? Unlike catecholamine and peptide hormones, steroid and thyroid hormones are lipid-soluble hormones and readily pass through the plasma membrane of a target cell into the cytoplasm. There they bind to specific intracellular receptor proteins, forming a complex that enters the nucleus and bind to specific regulatory sites on chromosomes. The binding alters the pattern of gene expression, initiating the transcription of some genes (DNA), while repressing the transcription of others. This results in the production of specific mRNA translation products, proteins and usually enzymes. The actions of lipid-soluble hormones are slower and last longer than the actions of water-soluble hormones. These cause physiological responses that are characteristic of the steroid hormones.
http://www.youtube.com/watch?v=oOj04WsU9ko
Examples of peptide hormones
Hormones of hypothalamus (releasing and inhibitory factors), structure, mechanism of action.
Hypothalamus has the wide anatomic links with other parts of the brain. Therefore in different mental disorders there is the change of secretion of hypothalamus hormones.
http://www.youtube.com/watch?v=hLeBNyB1qKU&feature=related
Two groups of hormones are produced by hypothalamus corresponding to the anterior and posterior pituitary.
Hypothalamus and posterior pituitary. 3 peptides are synthesized in the hypothalamus that pass to the posterior pituitary along axons where they are accumulated: oxytocin, vasopressin (antidiuretic hormone) and neurophysin. The later binds the oxytocin and vasopressin and promotes their transportation to the pituitary.
Hypothalamus and anterior pituitary. Hypothalamus is connected with the anterior pituitary by the net of blood capillaries, so called hypothalamic portal system. Hypothalamus produces very active peptide compounds that pass via this portal system to anterior pituitary and stimulate or oppress the secretion of tropic hormones. Compounds stimulating the secretion are called releasing factors. 7 releasing factors are known according to the amount of tropic hormones of anterior pituitary:
- corticotropin-releasing factor
- thyrotropin-releasing factors
- somatotropin-releasing factors
- follicletropin-releasing factor
- luteotropin-releasing factor
- prolactotropin-releasing factor
- melanotropin-releasing factor.
Hypothalamus also secretes substances called inhibitory factors or statins, which can inhibit release of the some pituitary hormones. 3 inhibitory factors are known today:
- somatostatin
- prolactostatin
- melanostatin.
Releasing and inhibitory factors are produced in only minute amounts.
http://www.youtube.com/watch?v=-UaSfYKsFh0
Hormones of pituitury, structure, mechanism of action.
Tropic hormones are produced by the anterior pituitary. Usually tropic hormones not directly regulate the metabolism but act on the peripheral endocrine glands.
|
|
Somatotropic hormone (STH, growth hormone)
Chemical structure: simple protein
The intensity of secretion is regulated by the relationship between the somatotropic-releasing factor and somatostatin.
The main function of somatotropic hormone - stimulation of growth. Hormone is necessary for the bone tissue formation, for the muscle tissue growth, for the formation of peculiarities of men and women body.
Somatotropic hormone can act both directly on the metabolism and indirectly stimulating the synthesis of somatomedines (specific protein growth factors which are synthesized in liver).
The effect of somatotropic hormone on:
- protein metabolism: stimulates the passing of amino acids into the cells;
activates the synthesis of proteins, DNA, RNA.
- carbohydrate metabolism: activates the insulinase of liver;
inhibits the conversion of lipids to carbohydrates;
activates the exit of glucose from liver;
inhibits the entry of glucose into the cells.
- lipid metabolism: stimulates lipolisis;
stimulates the oxidation of fatty acids.
The deficiency of somatotropic hormone in children age causes nanism. Nanism - proportional underdevelopment of all body.
The deficiency of somatotropic hormone in adult persons hasn’t clinical symptoms. The excess of somatotropic hormone in children age causes gigantism.
The excess of somatotropic hormone in adult persons causes acromegalia (disproportional development of the separate body parts).
http://www.youtube.com/watch?v=VX2wgM4kUfM
Thyrotropic hormone (TTH).
Chemical structure: glicoprotein.
This hormone is necessary for the normal functions of thyroid glands.
Thyrotropic hormone promotes:
- accumulation of iodine in thyroid;
- including of iodine into the tyrosine;
- synthesis of thyroxine and triiodothyronine.
Adrenocorticotropic hormone (ACTH).
Chemical structure: polipeptide.
This hormone is necessary for the normal functions of adrenal cortex. It enhances the formation of steroid hormones and their secretion into the blood.
ACTH has also the melanocyte-stimulating activity.
Excessive secretion of ACTH causes the Icenko-Kushing disease (symptoms of hypercorticism, hyperpigmentation).
As you know, Cushing’s is a rarely diagnosed endocrine disorder characterized by hypercortisolism. Cortisol is a hormone produced by the adrenal glands and is vital to regulate the body’s cardivoascular functions and metabolism, to boost the immune system and to fight inflammation. But its most important job is to help the body to respond to stress.
The adrenal glands release cortisol in response to stress, so atheletes, women experiencing pregnancy, and those suffering from alcoholism, panic disorders and malnutrition naturally have higher-than-normal levels of cortisol.
People with Cushing’s Syndrome live life with too much cortisol for their bodies as a result of a hormone-secreting tumor. Mine is located in the pituitary gland. Endogenous hypercortisolism leaves the body in a constant state of “fight or flight,” which ravages the body and tears down the body’s major systems including cardivascular, musculo-skeletal, endocrine, etc.
Symptoms vary, but the most common symptoms include rapid, unexplained weight gain in the upper body with increased fat around the neck and face (“moon facies”); buffalo hump; facial flushing/plethora; muscle wasting in the arms and legs; purplish striae (stretch marks) on the abdomen, thighs, buttocks, arms and breasts; poor wound healing and bruising; severe fatigue; depression, anxiety disorders and emotional lability; cognitive difficulties; sleep disorders due to abnormally high nighttime cortisol production; high blood pressure and high blood sugar/diabetes; edema; vision problems; premature osteoperosis; and, in women, signs of hyperandrogenism such as menstrual irregularities, infertility, hirsutism, male-patterned balding and steroid-induced acne.
Most people with Cushing’s long for the ability to do simple things,
like walk a flight of stairs without having to sit for half an hour afterwards,
or vacuum the house or even unload a dishwasher.
One of the worst parts about this disease is the crushing fatigue and muscle wasting/weakness, which accompanies hypercortisolism. Not only do we become socially isolated because of the virilzing effects of an endocrine tumor, which drastically alters our appearance, but we no longer feel like ourselves with regard to energy. We would love to take a long bike ride, run three miles or go shopping like we used to — activities, which we took for granted before the disease struck. Those activities are sadly impossible at times for those with advanced stages of the disease.
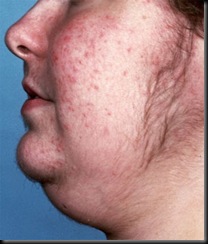
A patient with Cushing syndrome showing signs of acne and hirsuitism
Moon face in patienr with Cushing syndrome
Widened purple striae in a patient with Cushing's
Gonadotropic hormones.
Follicle stimulating hormone (FSH).
Chemical structure: glycoprotein.
Function: stimulates the function of follicles (oogenesis) in women and spermatogenesis in men.
FSH
(follicle stimulating hormone) regulates the development, growth, pubertal
maturation, and reproductive processes of the body
In both males and females, FSH stimulates the
maturation of germ cells.
In males, FSH induces sertoli cells to secrete inhibin and stimulates the
formation of sertoli-sertoli tight junctions (zonula occludens).
In females, FSH initiates follicular growth, specifically affecting granulosa cells.
With the concomitant rise in inhibin B, FSH levels then decline in the late
follicular phase. This seems to be critical in selecting only the most advanced
follicle to proceed to ovulation. At the end of the luteal phase, there is a
slight rise in FSH that seems to be of importance to start the next ovulatory
cycle.
Luteinizing hormone (LH).
Chemical structure: glycoprotein.
Function: stimulates the formation of yellow body in women and testosterone secretion in men.
In
both males and females, (LH) Luteinising hormone is essential for reproduction.
In females, at the time of menstruation, FSH initiates follicular growth,
specifically affecting granulosa cells. With the rise in estrogens, LH
receptors are also expressed on the maturing follicle that produces an
increasing amount of estradiol. Eventually at the time of the maturation of the
follicle, the estrogen rise leads via the 48 hour period.
This 'LH surge' triggers ovulation thereby not only releasing the egg, but also initiating the conversion of the residual follicle into a corpus luteum that, in turn, produces progesterone to prepare the endometrium for a possible implantation. LH is necessary to maintain luteal function for the first two weeks. In case of a pregnancy luteal function will be further maintained by the action of hCG (a hormone very similar to LH) from the newly established pregnancy. LH supports thecal cells in the ovary that provide androgens and hormonal precursors for estradiol production.
In the male, LH acts upon the Leydig cells of the testis and is responsible for the production of testosterone, an androgen that exerts both endocrine activity and intratesticular activity on spermatogenesis.
Prolactin (PRL).

Chemical structure: protein.
Functions: - stimulates lactation;
- stimulates function of yellow body (secretion of progesterone);
- promotes formation of mother instinct;
- stimulates the formation of prostate glandular tissue in men.
Lipotropic hormone.
Chemical structure: protein.
Functions: - stimulates the mobilization of lipids from depot;
- decreases the Ca amount in blood;
- has the melanocyte-stimulating activity.
|
|
|
Melanocyte and melanin. |

Posterior pituitary.
Vasopressin (Antidiuretic Hormone)
Chemical structure: peptide.
Functions: - activates the hyaluronidase. This enzyme splits the hyaluronic acid. The permeability of membranes is increased and reabsorption of water in kidneys is increased too. As result the day diuresis is decreased;
- narrows arterioles and capillaries and increases the blood pressure.
-
-
AVP has two principle sites of action: the kidney and blood vessels.
- The primary function of AVP in the body is to regulate extracellular fluid volume by affecting renal handling of water, although it is also a vasoconstrictor and pressor agent (hence, the name "vasopressin"). AVP acts on renal collecting ducts via V2 receptors to increase water permeability (cAMP-dependent mechanism), which leads to decreased urine formation (hence, the antidiuretic action of "antidiuretic hormone"). This increases blood volume, cardiac output and arterial pressure.
- A secondary function of AVP is vasoconstriction. AVP binds to V1 receptors on vascular smooth muscle to cause vasoconstriction via the IP3 signal transduction pathway, which increases arterial pressure; however, the normal physiological concentrations of AVP are below its vasoactive range. Studies have shown, nevertheless, that in severe hypovolemic shock, when AVP release is very high, AVP does contribute to the compensatory increase in systemic vascular resistance.
The deficiency of vasopressin in organism causes diabetes insipidus. Clinical symptoms - poliuria, dehydration of the organism, low density of the urine.
Diabetes insipidus results in excessive thirst and urination. The reason is problems with a particular hormone or its receptor. Diabetes insipidus increases the risk for dehydration.
What Is Diabetes Insipidus?
Diabetes insipidus is caused by problems related to a hormone called antidiuretic hormone or its receptor. Antidiuretic hormone (ADH) is produced in a part of the brain called the hypothalamus. It's stored in the brain's pituitary gland. Release of ADH causes the kidneys to hold onto water, which makes urine more concentrated.
Normally, if we are thirsty or slightly dehydrated, ADH levels rise. The kidneys reabsorb more water and excrete concentrated urine. If, on the other hand, we chugged a half-gallon of water (don't try this at home), ADH levels would fall. Clear, dilute urine would pass. Diabetes insipidus can be caused by either of two problems with ADH. One is too little ADH is produced. When that's the case, the condition is called central diabetes insipidus. The other is there's enough ADH produced, but the kidneys can't respond to it. That condition is known as nephrogenic diabetes insipidus.
In either form of diabetes insipidus, the result is the same. The kidneys can't do their job of conserving water. Even when a person with diabetes insipidus is dehydrated, the kidneys will excrete abundant, dilute urine. This inability of the kidneys to conserve water leads to the symptoms of diabetes insipidus:
- Excessive thirst
- Excessive urine production (polyuria)

In some people, these symptoms can become extreme, causing dehydration.
Excessive fluid losses can also cause electrolyte imbalances. Possible symptoms include:
- Unexplained weakness
- Lethargy
- Muscle pains
- Irritability
But why "insipidus?" People with diabetes insipidus aren't insipid, but their urine is. Insipid can mean dull or lacking flavor. Believe it or not, doctors long ago would taste urine to detect illness. Unlike diabetes mellitus, which results in sweet tasting urine, diabetes insipidus creates watery, flavor-free urine.
Oxytocin.
Chemical structure: peptide.
Functions: stimulates the contraction of smooth muscles, especially the muscles of uterus and muscle fibres of alveoluses of mammas.
Oxytocin is used for delivery stimulation, for stop of bleeding after delivery, for stimulation of lactation.
Numerically the largest group of signaling substances, these arise by protein biosynthesis. The smallest peptide hormone, thyroliberin (362 Da), is a tripeptide. Proteohormones can reach masses of more than 20 kDa—e. g., thyrotropin (28 kDa). Similarities in the primary structures of many peptide hormones and proteohormones show that they are related to one another. They probably arose from common predecessors in the course of evolution. Thyroliberin (thyrotropin-releasing hormone, TRH) is one of the neurohormones of the hypothalamus. It stimulates pituitary gland cells to secrete thyrotropin (TSH). TRH consists of three amino acids, which are modified in characteristic ways.
Thyrotropin (thyroid-stimulating hormone, TSH) and the related hormones lutropin (luteinizing hormone, LH) and follitropin (follicle-stimulating hormone, FSH) originate in the adenohypophysis. They are all dimeric glycoproteins with masses of around 28 kDa. Thyrotropin stimulates the synthesis and secretion of thyroxin by the thyroid gland.
Hormones of pancreas, structure, mechanism of action
Insulin is produced and released by the B cells of the pancreas and is released when the glucose level rises. Insulin reduces the blood sugar level by promoting processes that consume glucose— e. g., glycolysis, glycogen synthesis, and conversion of glucose into fatty acids. By contrast, it inhibits gluconeogenesis and glycogen degradation. Insulin causes cells in the liver, skeletal muscles, and fat tissue to absorb glucose from the blood. In the liver and skeletal muscles, glucose is stored as glycogen, and in fat cells (adipocytes) it is stored as triglycerides.
Insulin stops the use of fat as an energy source by inhibiting the release of glucagon. With the exception of the metabolic disorder diabetes mellitus and metabolic syndrome, insulin is provided within the body in a constant proportion to remove excess glucose from the blood, which otherwise would be toxic. When blood glucose levels fall below a certain level, the body begins to use stored sugar as an energy source through glycogenolysis, which breaks down the glycogen stored in the liver and muscles into glucose, which can then be utilized as an energy source. As a central metabolic control mechanism, its status is also used as a control signal to other body systems (such as amino acid uptake by body cells). In addition, it has several other anabolic effects throughout the body.
When control of insulin levels fails, diabetes mellitus can result. As a consequence, insulin is used medically to treat some forms of diabetes mellitus. Patients with type 1 diabetes depend on external insulin (most commonly injected subcutaneously) for their survival because the hormone is no longer produced internally.[2] Patients with type 2 diabetes are often insulin resistant and, because of such resistance, may suffer from a "relative" insulin deficiency. Some patients with type 2 diabetes may eventually require insulin if other medications fail to control blood glucose levels adequately. Over 40% of those with Type 2 diabetes require insulin as part of their diabetes management plan.
The human insulin protein is composed of 51 amino acids, and has a molecular weight of 5808 Da. It is a dimer of an A-chain and a B-chain, which are linked together by disulfide bonds.
http://www.youtube.com/watch?v=V1LjRi8Nvv4
http://www.youtube.com/watch?v=VLiTbb6MaEU&NR=1
http://www.youtube.com/watch?v=ZsTSoLhl3Y4&feature=related
http://www.youtube.com/watch?v=nBJN7DH83HA&feature=related
Glucagon,
a peptide of 29 amino acids, is a product of the A
cells of the pancreas. It is the antagonist of insulin and, like insulin,
mainly influences carbohydrate and lipid metabolism. Its effects are each
opposite to those of insulin. Glucagon mainly acts via the second messenger
Glucagon is a linear peptide of 29 amino acids. Its primary sequence is almost perfectly conserved among vertebrates, and it is structurally related to the secretin family of peptide hormones. Glucagon is synthesized as proglucagon and proteolytically processed to yield glucagon within alpha cells of the pancreatic islets. Proglucagon is also expressed within the intestinal tract, where it is processed not into glucagon, but to a family of glucagon-like peptides (enteroglucagon).
Physiologic Effects of Glucagon
The major effect of glucagon is to stimulate an increase in blood concentration of glucose. As discussed previously, the brain in particular has an absolute dependence on glucose as a fuel, because neurons cannot utilize alternative energy sources like fatty acids to any significant extent. When blood levels of glucose begin to fall below the normal range, it is imperative to find and pump additional glucose into blood. Glucagon exerts control over two pivotal metabolic pathways within the liver, leading that organ to dispense glucose to the rest of the body:
- Glucagon stimulates breakdown of glycogen stored in the liver. When blood glucose levels are high, large amounts of glucose are taken up by the liver. Under the influence of insulin, much of this glucose is stored in the form of glycogen. Later, when blood glucose levels begin to fall, glucagon is secreted and acts on hepatocytes to activate the enzymes that depolymerize glycogen and release glucose.
- Glucagon activates hepatic gluconeogenesis. Gluconeogenesis is the pathway by which non-hexose substrates such as amino acids are converted to glucose. As such, it provides another source of glucose for blood. This is especially important in animals like cats and sheep that don't absorb much if any glucose from the intestine - in these species, activation of gluconeogenic enzymes is the chief mechanism by which glucagon does its job.
Glucagon also appears to have a minor effect of enhancing lipolysis of triglyceride in adipose tissue, which could be viewed as an addition means of conserving blood glucose by providing fatty acid fuel to most cells.
Control of Glucagon Secretion
Knowing that glucagon's major effect is to increase blood glucose levels, it makes sense that glucagon is secreted in response to hypoglycemia or low blood concentrations of glucose.
Two other conditions are known to trigger glucagon secretion:
- Elevated blood levels of amino acids, as would be seen after consumption of a protein-rich meal: In this situation, glucagon would foster conversion of excess amino acids to glucose by enhancing gluconeogenesis. Since high blood levels of amino acids also stimulate insulin release, this would be a situation in which both insulin and glucagon are active.
- Exercise: In this case, it is not clear whether the actual stimulus is exercise per se, or the accompanying exercise-induced depletion of glucose.
In terms of negative control, glucagon secretion is inhibited by high levels of blood glucose. It is not clear whether this reflects a direct effect of glucose on the alpha cell, or perhaps an effect of insulin, which is known to dampen glucagon release. Another hormone well known to inhibit glucagon secretion is somatostatin.
Disease States
Diseases associated with excessively high or low secretion of glucagon are rare. Cancers of alpha cells (glucagonomas) are one situation known to cause excessive glucagon secretion. These tumors typically lead to a wasting syndrome and, interestingly, rash and other skin lesions.
Although insulin deficiency is clearly the major defect in type 1 diabetes mellitus, there is considerable evidence that aberrant secretion of glucagon contributes to the metabolic derangements seen in this important disease. For example, many diabetic patients with hyperglycemia also have elevated blood concentrations of glucagon, but glucagon secretion is normally suppressed by elevated levels of blood glucose.
What is diabetes mellitus?
Diabetes is a disease of the pancreas, an organ behind your stomach that produces the hormone insulin. Insulin helps the body use food(glucose) for energy. When a person has diabetes, the pancreas either cannot produce enough insulin, or the body uses the insulin incorrectly, or both. Insulin works together with glucose in the bloodstream to help it enter the body’s cells to be burned for energy. If the insulin isn’t functioning properly, glucose cannot enter the cells. This causes glucose levels in the blood to rise, creating a condition of high blood sugar or hyperglycaemia which is the hallmark of diabetes, and leaving the cells without fuel. When blood glucose rises above a certain level, it spills over into the urine.
What are the common types of diabetes?
There are two common forms of diabetes: type 1 and type 2.
- Type 1(Insulin dependent): Type 1 diabetes occurs because the insulin-producing cells of the pancreas are damaged. In type 1 diabetes, the pancreas makes little or no insulin, so sugar cannot get into the body’s cells for use as energy. People with type 1 diabetes must use insulin injections to control their blood glucose. Type 1 is the most common form of diabetes in people under age 20, but it can occur at any age. Ten percent of people with diabetes are diagnosed with type 1.
- Type 2(Non-insulin dependent): In type 2 diabetes, the pancreas makes insulin, but it either doesn’t produce enough insulin or the insulin does not work properly. Type 2 diabetes may sometimes be controlled with a combination of diet, weight management and exercise. However, treatment also may include oral glucose-lowering medications or insulin injections.
Generally, type 2 diabetes is more common in people over age 40 who are overweight. However, the increased prevalence of obesity has increased the number of people under age 40 who are diagnosed with type 2 diabetes. Nine out of 10 people with diabetes have type 2.
What causes Diabetes Mellitus?
The following factors may increase your chance of getting diabetes:
- Family history of diabetes or inherited tendency
- African-American, Hispanic or Native American race or ethnic background
- Obesity (being 20 percent or more over your desired body weight)
- Physical stress (such as surgery or illness)
- Use of certain medications
- Injury to pancreas (such as infection, tumor, surgery or accident)
- Autoimmune disease
- Hypertension
- Abnormal blood cholesterol or triglyceride levels
- Age (risk increases with age)
- Alcohol (risk increases with years of heavy alcohol use)
- Smoking
- Pregnancy (Pregnancy puts extra stress on a woman’s body which causes some women to develop diabetes. Blood sugar levels often return to normal after childbirth. Yet, women who develop diabetes during pregnancy have an increased chance of developing diabetes later in life.)
How is diabetes diagnosed?
The preferred method of diagnosing diabetes is the fasting blood sugar test (FBS). The FBS measures your blood glucose level after you have fasted (not eaten anything) for 10 to 12 hours.
Normal fasting blood glucose is between 70 and 100 mg/dl for people who do not have diabetes. The standard diagnosis of diabetes is made when:
- A patient has a fasting blood glucose level of 126 mg/dl or higher on two separate occasions; or
- A patient has a random blood glucose level of 200 mg/dl or greater and has common symptoms of diabetes, such as:
- – Increased thirst
- –Frequent urination
- –Increased hunger
- –Fatigue
–Blurred vision - –Weight loss
- On occasion, an oral glucose tolerance test may aid in the diagnosis of diabetes or an earlier abnormality that may become diabetes – called impaired glucose tolerance.
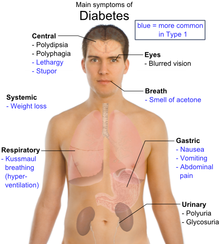
Other symptoms may include:
- Slow healing sores or cuts
- Itchy skin (usually in the vaginal or groin area); yeast infections
- Dry mouth
What are some of the long-term complications of diabetes?
Retinopathy (eye disease): All patients with diabetes should see an ophthalmologist (eye specialist) yearly for a dilated eye examination. Patients with known eye disease, symptoms of blurred vision in one eye or who have blind spots may need to see their ophthalmologist more frequently.
Nephropathy (kidney disease): Urine testing should be performed yearly. Regular blood pressure checks also are important because control of hypertension (high blood pressure) is essential in slowing kidney disease. Generally, blood pressure should be maintained less than 130/80 in adults. Persistent leg or feet swelling also may be a symptom of kidney disease and should be reported to your doctor.
Neuropathy (nerve disease): Numbness or tingling in your feet should be reported to your doctor at your regular visits. You should check your feet daily for redness, calluses, cracks or breakdown in skin tissue. If you notice these symptoms before scheduled visits, notify your doctor immediately.
Other long-term may complications include:
- Eye problems, such as glaucoma and cataracts
- Dental problems
- High blood pressure
- Heart disease
Because of the link between obesity and type 2 diabetes, you can do a great deal to reduce your chance of developing the disease by slimming down if you are overweight. This is especially true if diabetes runs in your family.
In fact,
studies have shown that exercise and a healthy diet can prevent the development
of type 2 diabetes in people with impaired glucose tolerance — a condition that
often develops prior to full-blown type 2 diabetes.
Medications have also been shown to provide similar benefit. Both diabetes
drugs metformin and Precose have been shown to prevent the onset of type 2
diabetes in people with this pre-diabetes condition.
In someone who already has diabetes, exercise and a nutritionally balanced diet
can greatly limit the effects of both types 1 and 2 diabetes on your body. In
diabetics, stopping smoking is one of the best ways to help prevent the
damaging effects of diabetes. If you smoke, quit; smoking dramatically
increases the risk of heart disease, particularly for people with diabetes.
http://www.youtube.com/watch?v=FEsTIOIufiQ
Hormones of adrenal glands.
Adrenal glands consist of two parts: external - cortex, internal - medulla.
http://www.youtube.com/watch?v=06jbq3bxKE0&feature=related
Epinephrine is a hormone synthesized in the adrenal glands from tyrosine. Its release is subject to neuronal control.This “emergency hormone” mainly acts on the blood vessels, heart, andmetabolism. It constricts the blood vessels and thereby increases blood pressure; it increases cardiac function; it promotes the degradation of glycogen into glucose in the liver and muscles; and it dilates the bronchia.
Each part secrets specific hormones.
Hormones of adrenal medulla – catecholamines (epinephrine, norepinephrine, dopamine).
Chemical structure - these hormones are derivatives of amino acid tyrosine.
Epinephrine, norepinephrine, dopamine exist in blood
in
Functions: causes very potent contraction of vessels and increase the blood pressure, increase a pulse rate. Epinephrine relaxes the smooth muscles of bronchi, intestine, promote the contraction of uterus smooth muscle. Epinephrine play a great role in stress reactions.
Catecholamine, any of various naturally occurring amines that function as neurotransmitters and hormones within the body. Catecholamines are characterized by a catechol group (a benzene ring with two hydroxyl groups) to which is attached an amine (nitrogen-containing) group. Among the catecholamines are dopamine, epinephrine (adrenaline), and norepinephrine (noradrenaline).
All catecholamines are synthesized from the amino acid l-tyrosine according to the following sequence: tyrosine → dopa (dihydroxyphenylalanine) → dopamine → norepinephrine (noradrenaline) → epinephrine (adrenaline). Catecholamines are synthesized in the brain, in the adrenal medulla, and by some sympathetic nerve fibres. The particular catecholamine that is synthesized by a nerve cell, or neuron, depends on which enzymes are present in that cell. For example, a neuron that has only the first two enzymes (tyrosine hydroxylase and dopa decarboxylase) used in the sequence will stop at the production of dopamine and is called a dopaminergic neuron (i.e., upon stimulation, it releases dopamine into the synapse). In the adrenal medulla the enzyme that catalyzes the transformation of norepinephrine to epinephrine is formed only in the presence of high local concentrations of glucocorticoids from the adjacent adrenal cortex; chromaffin cells in tissues outside the adrenal medulla are incapable of synthesizing epinephrine.
l-Dopa is well known for its role in the treatment of parkinsonism, but its biological importance lies in the fact that it is a precursor of dopamine, a neurotransmitter widely distributed in the central nervous system, including the basal ganglia of the brain (groups of nuclei within the cerebral hemispheres that collectively control muscle tone, inhibit movement, and control tremour). A deficiency of dopamine in these ganglia leads to parkinsonism, and this deficiency is at least partially alleviated by the administration of l-dopa.
Under ordinary circumstances, more epinephrine than norepinephrine is released from the adrenal medulla. In contrast, more norepinephrine is released from the sympathetic nervous system elsewhere in the body. In physiological terms, a major action of the hormones of the adrenal medulla and the sympathetic nervous system is to initiate a rapid, generalized fight-or-flight response. This response, which may be triggered by a fall in blood pressure or by pain, physical injury, abrupt emotional upset, or hypoglycemia, is characterized by an increased heart rate (tachycardia), anxiety, increased perspiration, tremour, and increased blood glucose concentrations (due to glycogenolysis, or breakdown of liver glycogen). These actions of catecholamines occur in concert with other neural or hormonal responses to stress, such as increases in adrenocorticotropic hormone (ACTH) and cortisol secretion.
Furthermore, the tissue responses to different catecholamines depend on the fact that there are two major types of adrenergic receptors (adrenoceptors) on the surface of target organs and tissues. The receptors are known as alpha-adrenergic and beta-adrenergic receptors, or alpha receptors and beta receptors, respectively. In general, activation of alpha-adrenergic receptors results in the constriction of blood vessels, contraction of uterine muscles, relaxation of intestinal muscles, and dilation of the pupils. Activation of beta-adrenergic receptors increases heart rate and stimulates cardiac contraction (thereby increasing cardiac output), dilates the bronchi (thereby increasing air flow into and out of the lungs), dilates the blood vessels, and relaxes the uterus. Drugs that block the activation of beta receptors (beta blockers), such as propranolol, are often given to patients with tachycardia, high blood pressure, or chest pain (angina pectoris). These drugs are contraindicated in patients with asthma because they worsen bronchial constriction.
Catecholamines play a key role in nutrient metabolism and the generation of body heat (thermogenesis). They stimulate not only oxygen consumption but also consumption of fuels, such as glucose and free fatty acids, thereby generating heat. They stimulate glycogenolysis and the breakdown of triglycerides, the stored form of fat, to free fatty acids (lipolysis). They also have a role in the regulation of secretion of multiple hormones. For example, dopamine inhibits prolactin secretion, norepinephrine stimulates gonadotropin-releasing hormone secretion, and epinephrine inhibits insulin secretion by the beta cells of the islets of Langerhans of the pancreas.
The effect of epinephrine on carbohydrate metabolism:
- activates the decomposition of glycogen in liver and muscles;
- activates the glycolysis, Krebs cycle and tissue respiration;
- causes the hyperglycemia.
The effect of epinephrine on protein metabolism:
- activates the protein decomposition.
The effect of epinephrine on lipid metabolism:
- activates the tissue lipase, mobilization of lipids and oxidation of fatty acids.
- Norepinephrine (NE) and epinephrine (E) are both sympathomimetic catecholamines that are synthesized in the adrenal medulla and terminals of sympathetic neurons. Along the metabolic pathway, NE is synthesized first -- and when an enzyme adds a methyl group, you have epinephrine. NE is the main catecholamine in peripheral tissue and sympathetic neurons. E is mostly made in the adrenal medulla.
-
http://www.youtube.com/watch?v=4g25d7_Afmc
Tissue hormones
Hormonoids (tissue hormones, histohormones) - organic trace substances produced by different cells of different tissues (not by specific glands) that regulate metabolism on the local level (some hormonoids are produced in the blood too (serotonin, acetylcholine).
In the organs, the hormones carry out physiological and biochemical regulatory functions. In contrast to endocrine hormones, tissue hormones are only active in the immediate vicinity of the cells that secrete them. The distinctions between hormones and other signaling substances (mediators, neurotransmitters, and growth factors) are fluid. Mediators is the term used for signaling substances that do not derive from special hormone- forming cells, but are formbymany cell types. Acetylcholine is a neurotransmitter. What that does is it releases chemicals into the brain and plays a role in normal brain functions such as sleep. Also, it deals with attention, learning, and memory skills. The mechanisms that the transmitter controls was a mystery until now. Scientist now know that acetycholine deals with communication between neurons. Located in the prefrontal cortex. This is the formula for acetylcholine.
When acetylcholine is released it binds to a specific reactor. Next it begins to start a molecule cascade. Which then triggers physiological alterations. Which deals with how prefrontal cortical neurons are “wired” together. This explains how acethlcholine is released into the brain. This process may actually have an effect in the formation of new associative memories. Most of this information can be found in the artical on acetylchonline. The Professor of Cellular Neuroscience, Zafar Bashir was the one who demonstrated how electron stimulation of the prefrontal cortex leads to the release of acetylcholine. Then Dr. Douglas Caruana carried out another experiment. He also found that acetylchonline when released into the prefrontal cortex it helps you remember things. But when to much has been released those memories start to be forgotten.
Just like the article said in the Journal of Neuroscience. Acetylcholine is a neurotransmitter which plays key roles in sleep and other normal functions. This is basically what acetylcholine does before of course we found out that to much of it is dangerous.
Gastrin is released in response to certain stimuli. These include:
- stomach distension
- vagal stimulation (mediated by the neurocrine bombesin, or GRP in humans)
- the presence of partially digested proteins especially amino acids
- hypercalcemia
Gastrin release is inhibited by:
- The presence of acid (primarily the secreted HCl) in the stomach (a case of negative feedback).
- Somatostatin also inhibits the release of gastrin, along with secretin, GIP (gastroinhibitory peptide), VIP (vasoactive intestinal peptide), glucagon and calcitonin.
Function
The presence of gastrin stimulates parietal cells of the stomach to secrete hydrochloric acid (HCl)/gastric acid. This is done both directly on the parietal cell and indirectly via binding onto CCK2/gastrin receptors on ECL cells in the stomach, which then responds by releasing histamine, which in turn acts in a paracrine manner on parietal cells stimulating them to secrete H+ ions. This is the major stimulus for acid secretion by parietal cells.
Along with the above mentioned function, gastrin has been shown to have additional functions as well:
- Stimulates parietal cell maturation and fundal growth.
- Causes chief cells to secrete pepsinogen, the zymogen (inactive) form of the digestive enzyme pepsin.
- Increases antral muscle mobility and promotes stomach contractions.
- Strengthens antral contractions against the pylorus, and relaxes the pyloric sphincter, which stimulates gastric emptying.
- Plays a role in the relaxation of the ileocecal valve.[8]
- Induces pancreatic secretions and gallbladder emptying.[9]
- Impacts lower esophageal sphincter (LES) tone, causing it to contract.
Heparin, a highly-sulfated glycosaminoglycan, is widely used as an injectable anticoagulant and has the highest negative charge density of any known biological molecule; it consists of a variably-sulfated repeating disaccharide unit: The most common disaccharide unit is composed of a 2-O-sulfated iduronic acid and 6-O-sulfated, N-sulfated glucosamine, IdoA-GlcN
Heparin is a
naturally-occurring anticoagulant produced by basophils and mast cells;
pharmaceutical grade heparin is derived from mucosal tissues of slaughtered
meat animals such as porcine intestine or bovine lung. Heparin allows the
body's natural clot lysis mechanisms to work normally to break down clots that have
already formed.
Heparin binds to the enzyme inhibitor antithrombin (AT) causing a conformational change that results in its activation through an increase in the flexibility of its reactive site loop. The activated AT then inactivates thrombin and other proteases involved in blood clotting, most notably factor Xa. The rate of inactivation of these proteases by AT can increase by up to 1000-fold due to the binding of heparin.
The conformational change inAT on heparin-binding
mediates its inhibition of factor Xa. For thrombin inhibition however, thrombin
must also bind to the heparin polymer at a site proximal to the
pentasaccharide. The highly-negative charge density of heparin contributes to
its very strong electrostatic interaction with thrombin The
formation of a ternary complex between AT, thrombin,
and heparin results in the inactivation of thrombin. For this reason heparin's
activity against thrombin is size-dependent, the ternary complex requiring at
least 18 saccharide units for efficient formation. In contrast anti factor Xa
activity only requires the pentasaccharide binding site
This size difference has led to the development of low-molecular-weight heparins
and more recently to fondaparinux as
pharmaceutical anticoagulants. Low-molecular-weight heparins and fondaparinux
target anti-factor Xa activity rather than anti-thrombin (IIa) activity, with
the aim of facilitating a more subtle regulation of coagulation and an improved
therapeutic index
Secretin hormone production is stimulated by acid chyme entering the duodenum. This hormone stimulates the pancreas to release bicarbonate to neutralize the acid.
Secretin is a hormone that both controls the environment in the duodenum by regulating secretions of the stomach and pancreas, and regulates water homeostasis throughout the body. It is produced in the S cells of the duodenum, which are located in the crypts of Lieberkühn. In humans, the secretin peptide is encoded by the SCT gene. Secretin was also the first hormone to be identified.
Secretin regulates the pH within the duodenum by inhibiting gastric acid secretion by the parietal cells of the stomach, and by stimulating bicarbonate production by the centroacinar cells and intercalated ducts of the pancreas.
In 2007, secretin was discovered to play a role in osmoregulation by acting on the hypothalamus, pituitary, and kidney.
Secretin increases watery bicarbonate solution from pancreatic and bile duct epithelium. Pancreatic centroacinar cells have secretin receptors in their plasma membrane. As secretin binds to these receptors, it stimulates adenylate cyclase activity and converts ATP to cyclic AMP. Cyclic AMP acts as second messenger in intracellular signal transduction and leads to increase in release of watery carbonate. It is known to promote the normal growth and maintenance of the pancreas.
Secretin increases water and bicarbonate secretion from duodenal Brunner's glands to buffer the incoming protons of the acidic chyme. It also enhances the effects of cholecystokinin to induce the secretion of digestive enzymes and bile from pancreas and gallbladder, respectively.
It counteracts blood glucose concentration spikes by triggering increased insulin release from pancreas, following oral glucose intake.
Although secretin releases gastrin from gastrinomas, it inhibits gastrin release from the normal stomach. It reduces acid secretion from the stomach by inhibiting gastrin release from G cells. This helps neutralize the pH of the digestive products entering the duodenum from the stomach, as digestive enzymes from the pancreas (e.g., pancreatic amylase and pancreatic lipase) function optimally at slightly basic pH.
In addition, secretin stimulates pepsin secretion from chief cells, which can help break down proteins in food digestion. It stimulates release of glucagon, pancreatic polypeptide and somatostatin.
Histamine, an important mediator (local signaling substance) and neurotransmitter, is mainly stored in tissue mast cells and basophilic granulocytes in the blood. It is involved in inflammatory and allergic reactions. “Histamine liberators” such as tissue hormones, type E immunoglobulins (see p. 300), and drugs can release it. Histamine acts via various types of receptor. Binding to H1 receptors promotes contraction of smoothmuscle in the bronchia, and dilates the capillary vessels and increases their permeability. Via H2 receptors, histamine slows down the heart rate and promotes the formation of HCl in the gastric mucosa. In the brain, histamine acts as a neurotransmitter.
They have hormone-like effects in their immediate surroundings. Histamine and prostaglandins are important examples of these substances. Histamine is found in plant and animal tissue and is released from mast cells as part of an allergic reaction in humans. Release of histamine stimulates gastric secretion and causes dilation of capillaries, constriction of bronchial smooth muscle, and decreases blood pressure.
Histamines are released from mast cells as an allergic response to abnormal proteins found in the blood. The mast cells are found in connective tissue that contains numerous basophilic granules and releases substances such as heparin and histamine in response to injury or inflammation of body tissues.
How severe can histamine reactions be?
It has recently been discovered that histamines may play a much larger roll in human disease than once thought. In the past, histamine production was blamed on some very common allergic reactions such as hay fever, bee sting reactions, and anaphylactic shock.
In recent studies, histamine involvement in chronic inflammatory and degenerative diseases such as Lupus, Arthritis, Gulf War Syndrome, Fibromyalgia, Leaky Gut Syndrome, and some skin disorders like Psoriasis and obscure Rashes, has come to light as causes of chronic inflammatory responses to abnormal proteins in the blood of chronically ill patients!
Histamine is a biogenic amine involved in local immune responses as well as regulating physiological function in the gut and acting as a neurotransmitter. Histamine triggers the inflammatory response. As part of an immune response to foreign pathogens, histamine is produced by basophils and by mast cells found in nearby connective tissues. Histamine increases the permeability of the capillaries to white blood cells and other proteins, in order to allow them to engage foreign invaders in the affected tissues. It is found in virtually all animal body cells.
Histamine
forms colorless hygroscopic crystals that melt at
Histamine has two basic centres, namely the aliphatic amino group and whichever nitrogen atom of the imidazole ring does not already have a proton. Under physiological conditions, the aliphatic amino group (having a pKa around 9.4) will be protonated, whereas the second nitrogen of the imidazole ring (pKa ≈ 5.8) will not be protonated. Thus, histamine is normally protonated to a singly-charged cation. Histamine is derived from the decarboxylation of the amino acid histidine, a reaction catalyzed by the enzyme L-histidine decarboxylase. It is a hydrophilic vasoactive amine.
Once formed, histamine is either stored or rapidly inactivated. Histamine released into the synapses is broken down by acetaldehyde dehydrogenase. It is the deficiency of this enzyme that triggers an allergic reaction as histamines pool in the synapses. Histamine is broken down by histamine-N-methyltransferase and diamine oxidase. Some forms of foodborne disease, so-called "food poisonings," are due to conversion of histidine into histamine in spoiled food, such as fish.
Serotonin is a neurotransmitter
= Neurotransmitters are chemicals that are used to relay, amplify and modulate signals between a neuron and another cell - cell to cell communicators
= Serotonin is produced in the body from amino acids
= Serotonin taken orally does not pass into the serotonergic pathways of the central nervous system because it does not cross the blood-brain barrier. However, tryptophan and its metabolite 5-hydroxytryptophan (5-HTP), from which serotonin is synthesized, can and does cross the blood-brain barrier. These agents are available as dietary supplements and may be effective serotonergic agents
= Drugs may hinder the natural use/loss of serotonin but Drugs do not incease the supply of serotonin.
Serotonin or 5-hydroxytryptamine (5-HT) is a monoamine neurotransmitter. Biochemically derived from tryptophan, serotonin is primarily found in the gastrointestinal (GI) tract, platelets, and in the central nervous system (CNS) of animals including humans. It is popularly thought to be a contributor to feelings of well-being and happiness.
Approximately 90% of the human body's total serotonin is located in the enterochromaffin cells in the alimentary canal (gut), where it is used to regulate intestinal movements. The remainder is synthesized in serotonergic neurons of the CNS, where it has various functions. These include the regulation of mood, appetite, and sleep. Serotonin also has some cognitive functions, including memory and learning. Modulation of serotonin at synapses is thought to be a major action of several classes of pharmacological antidepressants.
Serotonin secreted from the enterochromaffin cells eventually finds its way out of tissues into the blood. There, it is actively taken up by blood platelets, which store it. When the platelets bind to a clot, they disgorge serotonin, where it serves as a vasoconstrictor and helps to regulate hemostasis and blood clotting. Serotonin also is a growth factor for some types of cells, which may give it a role in wound healing.
Investigation of thyroid hormones in the regulation of metabolism. Hormonal regulation of calsium and phosphorus homeostasis.
Hormones of thyroid and parathyroid glands
Thyroid synthesizes two kinds of hormones: iodine containing hormones and calcitonin.
Iodine containing hormones - thyroxine and triiodthyronine.
Thyroid hormone is produced by the thyroid gland, which consists of follicles in which thyroid hormone is synthesized through iodination of tyrosine residues in the glycoprotein thyroglobulin. Thyroid stimulating hormone (TSH), secreted by the anterior pituitary in response to feedback from circulating thyroid hormone, acts directly on the TSH receptor (TSH-R) expressed on the thyroid follicular cell basolateral membrane. TSH regulates iodide uptake mediated by the sodium/iodide symporter, followed by a series of steps necessary for normal thyroid hormone synthesis and secretion. Thyroid hormone is essential for normal development, growth, neural differentiation, and metabolic regulation in mammals.
The THs, T4 and the more potent T3, are
synthesized in the thyroid gland. Iodide is actively transported and
concentrated into the thyroid by
The secretion of THs requires endocytosis of the stored iodinated Tg from the apical surface of the thyroid follicular cell. The internalized Tg is incorporated in phagolysosomes and undergoes proteolytic digestion, recapture of MIT and DIT, and release of T4 and T3 into the circulation via the basal surface. The majority of released TH is in the form of T4, as total serum T4 is 40-fold higher than serum T3 (90 vs. 2 nM). Only 0.03% of the total serum T4 is free (unbound), with the remainder bound to carrier proteins such as thyroxine binding globulin (TBG), albumin, and thyroid binding prealbumin. Approximately 0.3% of the total serum T3 is free, with the remainder bound to TBG and albumin. It is the free TH that enters target cells and generates a biological response.
The major pathway for the production of T3 is via 5′-deiodination of the outer ring of T4 by deiodinases and accounts for the majority of the circulating T3. Type I deioidinase is found in peripheral tissues such as liver and kidney and is responsible for the conversion of the majority of T4 to T3 in circulation. Type II deiodinase is found in brain, pituitary, and brown adipose tissue and primarily converts T4 to T3for intracellular use. These deiodinases recently have been cloned and demonstrated to be selenoproteins. 5′-Deiodination by type I deiodinase and type III deioidinase, which is found primarily in placenta, brain, and skin, leads to the generation of rT3, the key step in the inactivation of TH. rT3 and T3 can be further deiodinated in the liver and are sulfo- and glucuronide-conjugated before excretion in the bile. There also is an enterohepatic circulation of TH as intestinal flora deconjugates some of these compounds and promotes the reuptake of TH.
Although THs may exert their effects on a number of intracellular loci, their primary effect is on the transcriptional regulation of target genes. Early studies showed that the effects of THs at the genomic level are mediated by nuclear TRs, which are intimately associated with chromatin and bind TH with high affinity and specificity. Similar to steroid hormones that also bind to nuclear receptors, TH enters the cell and proceeds to the nucleus. It then binds to TRs, which may already be prebound to TREs located in promoter regions of target genes. The formation of ligand-bound TR complexes that are also bound to TREs is the critical first step in the positive or negative regulation of target genes and the subsequent regulation of protein synthesis. Given their abilities to bind both ligand and DNA as well as their ability to regulate transcription, TRs can be regarded as ligand-regulatable transcription factors.
Metabolism: Thyroid hormones stimulate diverse metabolic activities most tissues, leading to an increase in basal metabolic rate. One consequence of this activity is to increase body heat production, which seems to result, at least in part, from increased oxygen consumption and rates of ATP hydrolysis. By way of analogy, the action of thyroid hormones is akin to blowing on a smouldering fire. A few examples of specific metabolic effects of thyroid hormones include:
Lipid metabolism: Increased thyroid hormone levels stimulate fat mobilization, leading to increased concentrations of fatty acids in plasma. They also enhance oxidation of fatty acids in many tissues. Finally, plasma concentrations of cholesterol and triglycerides are inversely correlated with thyroid hormone levels - one diagnostic indiction of hypothyroidism is increased blood cholesterol concentration.
Carbohydrate metabolism: Thyroid hormones stimulate almost all aspects of carbohydrate metabolism, including enhancement of insulin-dependent entry of glucose into cells and increased gluconeogenesis and glycogenolysis to generate free glucose.
Protein metabolism: in normal concentration stimulate the synthesis of proteins and nucleic acids; in excessive concentration activate the catabolic processes.
Growth: Thyroid hormones are clearly necessary for normal growth in children and young animals, as evidenced by the growth-retardation observed in thyroid deficiency. Not surprisingly, the growth-promoting effect of thyroid hormones is intimately intertwined with that of growth hormone, a clear indiction that complex physiologic processes like growth depend upon multiple endocrine controls.
Development: Of critical importance in mammals is the fact that normal levels of thyroid hormone are essential to the development of the fetal and neonatal brain.
Other Effects: As mentioned above, there do not seem to be organs and tissues that are not affected by thyroid hormones. A few additional, well-documented effects of thyroid hormones include:
Cardiovascular system: Thyroid hormones increases heart rate, cardiac contractility and cardiac output. They also promote vasodilation, which leads to enhanced blood flow to many organs.
Central nervous system: Both decreased and increased concentrations of thyroid hormones lead to alterations in mental state. Too little thyroid hormone, and the individual tends to feel mentally sluggish, while too much induces anxiety and nervousness.
Reproductive system:
Thyroid Disease States
Disease is associated with both inadequate production and overproduction of thyroid hormones. Both types of disease are relatively common afflictions of man and animals.
Hypothyroidism is the result from any condition that results in thyroid hormone deficiency. Two well-known examples include:
Iodine deficiency: Iodide is absolutely necessary for production of thyroid hormones; without adequate iodine intake, thyroid hormones cannot be synthesized. Historically, this problem was seen particularly in areas with iodine-deficient soils, and frank iodine deficiency has been virtually eliminated by iodine supplementation of salt.
Primary thyroid disease: Inflammatory diseases of the thyroid that destroy parts of the gland are clearly an important cause of hypothyroidism.
Common symptoms of hypothyroidism arising after early childhood include lethargy, fatigue, cold-intolerance, weakness, hair loss and reproductive failure. If these signs are severe, the clinical condition is called myxedema. In the case of iodide deficiency, the thyroid becomes inordinantly large and is called a goiter.
About 95 percent of the active thyroid hormone is thyroxine, and most of the remaining 5 percent is triiodothyronine. Both of these require iodine for their synthesis. Thyroid hormone secretion is regulated by a negative feedback mechanism that involves the amount of circulating hormone, hypothalamus, and adenohypophysis.
If there is an iodine deficiency, the thyroid cannot make sufficient hormone. This stimulates the anterior pituitary to secrete thyroid-stimulating hormone, which causes the thyroid gland to increase in size in a vain attempt to produce more hormones. But it cannot produce more hormones because it does not have the necessary raw material, iodine. This type of thyroid enlargement is called simple goiter or iodine deficiency goiter.
Calcitonin is secreted by the parafollicular cells of the thyroid gland. This hormone opposes the action of the parathyroid glands by reducing the calcium level in the blood. If blood calcium becomes too high, calcitonin is secreted until calcium ion levels decrease to normal.
The most severe and devestating form of hypothyroidism is seen in young children with congenital thyroid deficiency. If that condition is not corrected by supplemental therapy soon after birth, the child will suffer from cretinism, a form of irreversible growth and mental retardation.
Congenital
hypothyroidism can
be endemic, genetic, or sporadic. If untreated, it results in mild to severe
impairment of both physical and mental growth and development. Poor
length growth is apparent as early as the first year of life. Adult stature
without treatment ranges from 1 to
Sporadic and genetic cretinism results from abnormal development or function of the foetal thyroid gland. This type of cretinism has been almost completely eliminated in developed countries by early diagnosis by newborn screening schemes followed by lifelong treatment with thyroxine (T4).
Thyroxine must be dosed as tablets only, even to newborns, as the liquid oral suspensions and compounded forms cannot be depended on for reliable dosing. In the case of dosing infants, the T4 tablets are generally crushed and mixed with breast milk, formula milk or water. If the medication is mixed with formulas containing iron or soya products, larger doses may be required, as these substances may alter the absorption of thyroid hormone from the gut. Frequent monitoring (every 2–3 weeks during the first months of life) is recommended to ensure that infants with congenital hypothyroidism remain within the high end of normal range, or euthyroid.
Cretinism arises from a diet deficient in iodine. It has affected many people worldwide and continues to be a major public health problem in many countries. Iodine is an essential trace element, necessary primarily for the synthesis of thyroid hormones. Iodine deficiency is the most common preventable cause of brain damage worldwide. Although iodine is found in many foods, it is not universally present in all soils in adequate amounts. Most iodine, in iodide form, is in the oceans where the iodide ions oxidize to elemental iodine, which then enters the atmosphere and falls to earth as rain, introducing iodine to soils. Earth deficient in iodine is most common inland and in mountainous areas and areas of frequent flooding, but can also occur in coastal regions owing to past glaciation, and leaching by snow, water and heavy rainfall, which removes iodine from the soil.[8] Plants and animals grown in iodine deficient soils are correspondingly deficient. Populations living in those areas without outside food sources are most at risk of iodine deficiency.
Diseases.
Calcitonin is synthesized by the parafollicle cells of thyroid.
Functions: - promotes the transition of calcium from blood in bones;
- inhibits the reabsorption of phosphorus in kidneys.
Thus, calcitonin decreases the Ca and P contents in blood.
Parathyroid hormone. Chemical structure: protein.
1. promotes the transition of calcium from bones to blood;
2. promotes the absorption of Ca in the intestine;
3. inhibits the reabsorption of phosphorus in kidneys.
Thus, parathyroid hormone increases the Ca amount in blood and decreases the P amount in blood.
Body Distribution of Calcium and Phosphate
There are three major pools of calcium in the body:
Calcium in blood and extracellular fluid:
Fluxes of Calcium and Phosphate
· Inhibition of bone resorption, which would minimize fluxes of calcium from bone into blood.
http://www.youtube.com/watch?v=JwPVibQ6_3Y&feature=related
http://www.youtube.com/watch?v=n7vybcT9_F4
· Stimulates production of the biologically-active form of vitamin D within the kidney.
Adrenal glands consist of two parts: external - cortex, internal - medulla.
Each part secrets specific hormones.
Hormones synthesized in adrenal cortex are named corticosteroids.
Mechanism of steroid hormones action (permeating into the cells):
http://www.youtube.com/watch?v=oOj04WsU9ko
http://www.youtube.com/watch?v=0ss8YIoKw0g
Structure of Intracellular Receptors
- The amino-terminus: In most cases, this region is involved in activating or stimulating transcription by interacting with other components of the transcriptional machinery. The sequence is highly variable among different receptors.
- DNA binding domain: Amino acids in this region are responsible for binding of the receptor to specific sequences of DNA.
- The carboxy-terminus or ligand-binding domain: This is the region that binds hormone.
Hormone-Receptor Binding and Interactions with DNA
- Receptor activation is the term used to describe conformational changes in the receptor induced by binding hormone. The major consequence of activation is that the receptor becomes competent to bind DNA.
- Activated receptors bind to "hormone response elements", which are short specific sequences of DNA which are located in promoters of hormone-responsive genes. In most cases, hormone-receptor complexes bind DNA in pairs, as shown in the figure below.
- Transcription from those genes to which the receptor is bound is affected. Most commonly, receptor binding stimulates transcription. The hormone-receptor complex thus functions as a transcription factor.
The synthesis of corticosteroids is regulated by ACTH.
In the blood corticosteroids are connected with proteins and transported to different organs.
Time half-life for corticosteroids is about 1 hour.
Ways of metabolism of corticosteroids:
2. Oxidation of 21-st carbon atom.
4. Corticosteroids can be excreted by kidneys in native structure.
http://www.youtube.com/watch?v=0ss8YIoKw0g
The effect of glucocorticoids on protein metabolism:
2. stimulate the activity of aminotransferases;
3. activate the synthesis of urea.
The effect of glucocorticoids on carbohydrate metabolism:
1. activate the gluconeogenesis;
2. inhibit the activity of hexokinase;
3. activate the glycogen synthesis in liver.
Glucocorticoids causes the hyperglycemia.
The effect of glucocorticoids on lipid metabolism:
1. promote the absorption of lipids in intestine;
3. activate the conversion of fatty acids in carbohydrates.
http://www.youtube.com/watch?v=ku-QJyQ0j7M&feature=related
· adults (8 A.M.): 6-28 mg/dL; adults (4 P.M.): 2-12 mg/dL
· child one to six years (8 A.M.): 3-21 mg/dL; child one to six years (4 P.M.): 3-10 mg/dL
· adolescent: 5-55 mg/24 hours
ACTH Dependent
(80%)
![]() Pituitary
Tumors (60%)
Pituitary
Tumors (60%)
ACTH
Independent (20%)
![]() Benign
Adrenal Tumors (adenoma) (25%)
Benign
Adrenal Tumors (adenoma) (25%)
![]() Malignant Adrenal
Tumors (adrenal cell carcinoma) (10%)
Malignant Adrenal
Tumors (adrenal cell carcinoma) (10%)
Treatment of Cushings Syndrome
![]() Functions of
mineralocorticoids.
Functions of
mineralocorticoids.
Secretion of mineralocorticoids is regulated by renin-angiotensine system
- activates the reabsorption of Na+, Cl- and water in kidney canaliculuses;
- promote the excretion of K+ by kidneys, skin and saliva.
Deficiency of corticosteroids causes Addison's disease.
http://www.youtube.com/watch?v=FK1pPqWMXjM
A rare disorder in which symptoms are caused by a deficiency of hydrocortisone (cortisol) and aldosterone, two corticosteroid hormones normally produced by a part of the adrenal glands called the adrenal cortex. Symptoms include weakness, tiredness, vague abdominal pain, weight loss, skin pigmentation and low blood pressure. Mineralocorticoids
Primary hyperaldosteronism has many causes, including adrenal hyperplasia and adrenal carcinoma.[2]
· Bilateral (micronodular) adrenal hyperplasia, 60%
· Adrenal (Conn's) adenoma, 40%
· Glucocorticoid-remediable hyperaldosteronism (dexamethasone-suppressible hyperaldosteronism), <1%
· rare forms, including disorders of the renin-angiotensin system, <1%
Male sex hormones are called androgens and female - estrogens.
Chemical structure - steroids.
Estrogens, in females, are produced primarily by the ovaries, and during pregnancy, the placenta. Follicle-stimulating hormone(FSH) stimulates the ovarian production of estrogens by the granulosa cells of the ovarian follicles and corpora lutea. Some estrogens are also produced in smaller amounts by other tissues such as the liver, adrenal glands, and the breasts. These secondary sources of estrogens are especially important in postmenopausal women. Fat cells produce estrogen as well.
· Promote formation of female secondary sex characteristics
· Stimulate endometrial growth
· Increase vaginal lubrication
· Maintenance of vessel and skin
· Reduce bone resorption, increase bone formation
· Increase hepatic production of binding proteins
· Increase circulating level of factors 2, 7, 9, 10, plasminogen
· Increase platelet adhesiveness
· Decrease LDL, fat deposition
· Salt (sodium) and water retention
· Increase cholesterol in bile
· Increase pheomelanin, reduce eumelanin
· Support hormone-sensitive breast cancers (see section below)
· Promotes lung function by supporting alveoli (in rodents but probably in humans).
· Progesterone decreases contractility of the uterine smooth muscle.
· A drop in progesterone levels is possibly one step that facilitates the onset of labor.
The fetus metabolizes placental progesterone in the production of adrenal steroids.
http://www.youtube.com/watch?v=nLmg4wSHdxQ&feature=fvwrel
Besides testosterone, other androgens include:
Effect of sex hormones on protein metabolism:
1. stimulate the processes of protein, DNA, RNA synthesis;
2. cause the positive nitrogenous equilibrium.
Effect of sex hormones on carbohydrate metabolism:
2. activate the synthesis of glycogen in liver.
Effect of sex hormones on lipid metabolism:
1. enhance the oxidation of lipids;
2. inhibit the synthesis of cholesterol.
Effect of sex hormones on energy metabolism:
- stimulate the Krebs cycle, tissue respiration and ATP production.
There are a variety of physiological effects including:
Basic principles of metabolism: catabolism, anabolism. Common pathways of proteins, carbohydrates and lipids transformation.
Investigation of Krebs cycle functioning.
Metabolism is the set of life-sustaining chemical transformations within the cells of living organisms. These enzyme-catalyzed reactions allow organisms to grow and reproduce, maintain their structures, and respond to their environments. The word metabolism can also refer to all chemical reactions that occur in living organisms, including digestion and the transport of substances into and between different cells, in which case the set of reactions within the cells is called intermediary metabolism or intermediate metabolism.
The term metabolism is derived from the Greek – "Metabolismos" for "change", or "overthrow". The history of the scientific study of metabolism spans several centuries and has moved from examining whole animals in early studies, to examining individual metabolic reactions in modern biochemistry. The first controlled experiments in human metabolism were published by Santorio Santorioin 1614 in his book Ars de statica medicina. He described how he weighed himself before and after eating, sleep, working, sex, fasting, drinking, and excreting. He found that most of the food he took in was lost through what he called "insensible perspiration".
In these early studies, the mechanisms of these metabolic processes had not been identified and a vital force was thought to animate living tissue. In the 19th century, when studying the fermentation of sugar to alcohol by yeast, Louis Pasteur concluded that fermentation was catalyzed by substances within the yeast cells he called "ferments". He wrote that "alcoholic fermentation is an act correlated with the life and organization of the yeast cells, not with the death or putrefaction of the cells."] This discovery, along with the publication by Friedrich Wöhler in 1828 of the chemical synthesis of urea, notable for being the first organic compound prepared from wholly inorganic precursors, proved that the organic compounds and chemical reactions found in cells were no different in principle than any other part of chemistry.
It was the discovery of enzymes at the beginning of the 20th century by Eduard Buchner that separated the study of the chemical reactions of metabolism from the biological study of cells, and marked the beginnings of biochemistry. The mass of biochemical knowledge grew rapidly throughout the early 20th century. One of the most prolific of these modern biochemists wasHans Krebs who made huge contributions to the study of metabolism. He discovered the urea cycle and later, working with Hans Kornberg, the citric acid cycle and the glyoxylate cycle. Modern biochemical research has been greatly aided by the development of new techniques such as chromatography, X-ray diffraction, NMR spectroscopy, radioisotopic labelling, electron microscopy andmolecular dynamics simulations. These techniques have allowed the discovery and detailed analysis of the many molecules and metabolic pathways in cells.
Metabolism is a term that is used to describe all chemical reactions involved in maintaining the living state of the cells and the organism. Metabolism can be conveniently divided into two categories:
· Catabolism - the breakdown of molecules to obtain energy
· Anabolism - the synthesis of all compounds needed by the cells
Anabolism is the set of constructive metabolic processes where the energy released by catabolism is used to synthesize complex molecules. In general, the complex molecules that make up cellular structures are constructed step-by-step from small and simple precursors. Anabolism involves three basic stages. Firstly, the production of precursors such as amino acids, monosaccharides,isoprenoids and nucleotides, secondly, their activation into reactive forms using energy from ATP, and thirdly, the assembly of these precursors into complex molecules such as proteins, polysaccharides, lipids and nucleic acids.
Metabolism refers to the highly integrated network of chemical reactions by which living cells grow and sustain themselves. This network is composed of two major types of pathways: anabolism and catabolism. Anabolism uses energy stored in the form of adenosine triphosphate (ATP) to build larger molecules from smaller molecules. Catabolic reactions degrade larger molecules in order to produce ATP and raw materials for anabolic reactions.
Together, these two general metabolic networks have three major functions:
(1) to extract energy from nutrients or solar energy;
(2) to synthesize the building blocks that make up the large molecules of life: proteins, fats, carbohydrates, nucleic acids, and combinations of these substances;
(3) to synthesize and degrade molecules required for special functions in the cell.
These reactions are controlled by enzymes, protein catalysts that increase the speed of chemical reactions in the cell without themselves being changed. Each enzyme catalyzes a specific chemical reaction by acting on a specific substrate, or raw material. Each reaction is just one in a sequence of catalyticsteps known as metabolic pathways. These sequences may be composed of up to20 enzymes, each one creating a product that becomes the substrate--or raw material--for the subsequent enzyme. Often, an additional molecule called a coenzyme is required for the enzyme to function. For example, some coenzymes accept an electron that is released from the substrate during the enzymatic reaction. Most of the water-soluble vitamins of the B complex serve as coenzymes;riboflavin (Vitamin B2) for example, is a precursor of the coenzyme flavine adenine dinucleotide, while pantothenate is a component of coenzyme A, an important intermediate metabolite.
The series of products created by the sequential enzymatic steps of anabolismor catabolism are called metabolic intermediates, or metabolites. Each steprepresents a small change in the molecule, usually the removal, transfer, oraddition of a specific atom, molecule, or group of atoms that serves as a functional group, such as the amino groups (-NH2) of proteins.
Most such metabolic pathways are linear, that is, they begin with a specificsubstrate and end with a specific product. However, some pathways, such as the Krebs cycle, are cyclic. Often, metabolic pathways also have branches thatfeed into or out of them. The specific sequences of intermediates in the pathways of cell metabolism are called intermediary metabolism.
Among the many hundreds of chemical reactions there are only a few that are central to the activity of the cell, and these pathways are identical in mostforms of life.
All reactions of metabolism, however, are part of the overall goal of the organism to maintain its internal orderliness, whether that organism is a singlecelled protozoan or a human. Organisms maintain this orderliness by removingenergy from nutrients or sunlight and returning to their environment an equal amount of energy in a less useful form, mostly heat. This heat becomes dissipated throughout the rest of the organism's environment.
According to the first law of thermodynamics, in any physical or chemical change, the total amount of energy in the universe remains constant, that is, energy cannot be created or destroyed. Thus, when the energy stored in nutrientmolecules is released and captured in the form of ATP, some energy is lost as heat. But the total amount of energy is unchanged.
The second law of thermodynamics states that physical and chemical changes proceed in such a direction that useful energy undergoes irreversible degradation into a randomized form--entropy. The dissipation of energy during metabolism represents an increase in the randomness, or disorder, of the organism's environment. Because this disorder is irreversible, it provides the driving force and direction to all metabolic enzymatic reactions.
Even in the simplest cells, such as bacteria, there are at least a thousand such reactions. Regardless of the number, all cellular reactions can be classified as one of two types of metabolism: anabolism and catabolism. These reactions, while opposite in nature, are linked through the common bond of energy.Anabolism, or biosynthesis, is the synthetic phase of metabolism during which small building block molecules, or precursors, are built into large molecular components of cells, such as carbohydrates and proteins.
Catabolic reactions are used to capture and save energy from nutrients, as well as to degrade larger molecules into smaller, molecular raw materials for reuse by the cell. The energy is stored in the form of energy-rich ATP, whichpowers the reactions of anabolism. The useful energy of ATP is stored in theform of a high-energy bond between the second and third phosphate groups of ATP. The cell makes ATP by adding a phosphate group to the molecule adenosinediphosphate (ADP). Therefore, ATP is the major chemical link between the energy-yielding reactions of catabolism, and the energy-requiring reactions of anabolism.
In some cases, energy is also conserved as energy-rich hydrogen atoms in thecoenzyme nicotinamide adenine dinucleotide phosphate in the reduced form of NADPH. The NADPH can then be used as a source of high-energy hydrogen atoms during certain biosynthetic reactions of anabolism.
In addition to the obvious difference in the direction of their metabolic goals, anabolism and catabolism differ in other significant ways. For example, the various degradative pathways of catabolism are convergent. That is, many hundreds of different proteins, polysaccharides, and lipids are broken down into relatively few catabolic end products. The hundreds of anabolic pathways,however, are divergent. That is, the cell uses relatively few biosynthetic precursor molecules to synthesize a vast number of different proteins, polysaccharides, and lipids.
The opposing pathways of anabolism and catabolism may also use different reaction intermediates or different enzymatic reactions in some of the steps. Forexample, there are 11 enzymatic steps in the breakdown of glucose into pyruvic acid in the liver. But the liver uses only nine of those same steps in thesynthesis of glucose, replacing the other two steps with a different set ofenzyme-catalyzed reactions. This occurs because the pathway to degradation ofglucose releases energy, while the anabolic process of glucose synthesis requires energy. The two different reactions of anabolism are required to overcome the energy barrier that would otherwise prevent the synthesis of glucose.
Another reason for having slightly different pathways is that the corresponding anabolic and catabolic routes must be independently regulated. Otherwise,if the two phases of metabolism shared the exact pathway (only in reverse) aslowdown in the anabolic pathway would slow catabolism, and vice versa.
Some reactions can be either catabolic or anabolic, depending on the circumstances. Such reactions are called amphibolic reactions. Many of the reactions interconverting the “simple molecules” fall in this category.
Catabolic and anabolic pathways are interrelated in three ways:
Matter (catabolic pathways furnish the precursor compounds for anabolism. Energy (catabolic pathways furnish the energy to “drive” anabolism). Electrons (catabolic pathways furnish the reducing power for anabolism).
Linear pathways convert one compound through a series of intermediates to another compound. An example would be glycolysis, where glucose is converted to pyruvate.
Branched pathways may either be divergent (an intermediate can enter several linear pathways to different end products) or convergent (several precursors can give rise to a common intermediate). Biosynthesis of purines and of some amino acids are examples of divergent pathways. There is usually some regulation at the branch point. The conversion of various carbohydrates into the glycolytic pathway would be an example of convergent pathways.
In a cyclic pathway, intermediates are regenerated, and so some intermediates act in a catalytic fashion. In this illustration, the cyclic pathway carries out the net conversion of X to Z. The Tricarboxylic Acid Cycle is an example of a cyclic pathway.
A pool of compounds in equilibrium with each other provides the intermediates for converting compounds to a variety of products, depending on what is fed “into” the pool and what is “withdrawn” from the pool. The phosphogluconate pathway is an example of such a pool of intermediates. The pathway can convert glucose to CO2, hexoses to pentoses, pentoses to hexoses, pentoses to trioses, etc. depending on what the cell requires in a particular situation. NADPH as a source of reducing power for anabolic reactions is also a main product of the phosphogluconate pathway.
Organisms differ in how many of the molecules in their cells they can construct for themselves. Autotrophs such as plants can construct the complex organic molecules in cells such as polysaccharides and proteins from simple molecules like carbon dioxide and water. Heterotrophs, on the other hand, require a source of more complex substances, such as monosaccharides and amino acids, to produce these complex molecules. Organisms can be further classified by ultimate source of their energy: photoautotrophs and photoheterotrophs obtain energy from light, whereas chemoautotrophs and chemoheterotrophs obtain energy from inorganic oxidation reactions.
Metabolism is closely linked to nutrition and the availability of nutrients. Bioenergetics is a term which describes the biochemical or metabolic pathways by which the cell ultimately obtains energy. Energy formation is one of the vital components of metabolism.
The speed of metabolism, the metabolic rate, influences how much food an organism will require, and also affects how it is able to obtain that food.
A striking feature of metabolism is the similarity of the basic metabolic pathways and components between even vastly different species. For example, the set of carboxylic acids that are best known as the intermediates in the citric acid cycle are present in all known organisms, being found in species as diverse as the unicellular bacterium Escherichia coli and huge multicellular organisms.
Nutrition is the key to metabolism. The pathways of metabolism rely upon nutrients that they breakdown in order to produce energy. This energy in turn is required by the body to synthesize new proteins, nucleic acids (DNA, RNA) etc.
Nutrients in relation to metabolism encompass bodily requirement for various substances, individual functions in body, amount needed, level below which poor health results etc.
Essential nutrients supply energy (calories) and supply the necessary chemicals which the body itself cannot synthesize. Food provides a variety of substances that are essential for the building, upkeep, and repair of body tissues, and for the efficient functioning of the body.
The diet needs essential nutrients like carbon, hydrogen, oxygen, nitrogen, phosphorus, sulfur, and around 20 other inorganic elements. The major elements are supplied incarbohydrates, lipids, and protein. In addition, vitamins, minerals and water are necessary.
The fate of dietary components after digestion and absorption constitutes metabolism—the metabolic pathways taken by individual molecules, their interrelationships, and the mechanisms that regulate the flow of metabolites through the pathways. Metabolic pathways fall into three categories: (1) Anabolic pathways are those involved in the synthesis of compounds. Protein synthesis is such a pathway, as is the synthesis of fuel reserves of triacylglycerol and glycogen. Anabolic pathways are endergonic. (2) Catabolic pathways are involved in the breakdown of larger molecules, commonly involving oxidative reactions; they are exergonic, producing reducing equivalents and, mainly via the respiratory chain, ATP.
Amphibolic pathways occur at the “crossroads” of metabolism, acting as links between the anabolic and catabolic pathways, eg, the citric acid cycle.
A knowledge of normal metabolism is essential for an understanding of abnormalities underlying disease. Normal metabolism includes adaptation to periods of starvation, exercise, pregnancy, and lactation. Abnormal metabolism may result from nutritional deficiency, enzyme deficiency, abnormal secretion of hormones, or the actions of drugs and toxins. An important example of a metabolic disease is diabetes mellitus.
PATHWAYS THAT PROCESS THE MAJOR PRODUCTS OF DIGESTION
The nature of the diet sets the basic pattern of metabolism. There is a need to process the products of digestion of dietary carbohydrate, lipid, and protein. These are mainly glucose, fatty acids and glycerol, and amino acids, respectively. In ruminants (and to a lesser extent in other herbivores), dietary cellulose is fermented by symbiotic microorganisms to short-chain fatty acids (acetic, propionic, butyric), and metabolism in these animals is adapted to use these fatty acids as major substrates.
All the products of digestion are metabolized to a common product, acetyl-CoA, which is then oxidized by the citric acid cycle .
Carbohydrate Metabolism Is Centered on the Provision & Fate of Glucose
Glucose is metabolized to pyruvate by the pathway of glycolysis, which can occur anaerobically (in the absence of oxygen), when the end product is lactate. Aerobic tissues metabolize pyruvate to acetyl-CoA, which can enter the citric acid cycle for complete oxidation to CO2 and H2O, linked to the formation of ATP.
Glucose and its metabolites also take part in other processes. Examples: (1) Conversion to the storage polymer glycogen in skeletal muscle and liver. (2) The pentose phosphate pathway, an alternative to part of the pathway of glycolysis, is a source of reducing equivalents (NADPH) for biosynthesis and the source of ribose for nucleotide and nucleic acid synthesis. (3) Triose phosphate gives rise to the glycerol moiety of triacylglycerols. (4) Pyruvate and intermediates of the citric acid cycle provide the carbon skeletons for the synthesis of amino acids; and acetyl-CoA, the precursor of fatty acids and cholesterol (and hence of all steroids synthesized in the body). Gluconeogenesis is the process of forming glucose from noncarbohydrate precursors, eg, lactate, amino acids, and glycerol.
Foods supply carbohydrates in three forms: starch, sugar, and cellulose (fiber). Starches and sugars form major and essential sources of energy for humans. Fibers contribute to bulk in diet.
Body tissues depend on glucose for all activities. Carbohydrates and sugars yield glucose by digestion or metabolism.Most people consume around half of their diet as carbohydrates.
Lipid Metabolism Is Concerned Mainly With Fatty Acids & Cholesterol
The source of long-chain fatty acids is either dietary lipid or de novo synthesis from acetyl-CoA derived from carbohydrate. Fatty acids may be oxidized to acetyl- CoA (β-oxidation) or esterified with glycerol, forming triacylglycerol (fat) as the body’s main fuel reserve. Acetyl-CoA formed by β-oxidation may undergo several fates:
(1) As with acetyl-CoA arising from glycolysis, it is oxidized to CO2 + H2O via the citric acid cycle.
(2) It is the precursor for synthesis of cholesterol and other steroids.
(3) In the liver, it forms ketone bodies (acetone, acetoacetate, and 3 hydroxybutyrate) that are important fuels in prolonged starvation.
Fats are concentrated sources of energy. They produce twice as much energy as either carbohydrates or protein on a weight basis.
Carbohydrate catabolism is the breakdown of carbohydrates into smaller units. Carbohydrates are usually taken into cells once they have been digested intomonosaccharides. Once inside, the major route of breakdown is glycolysis, where sugars such as glucose and fructose are converted into pyruvate and some ATP is generated.
Pyruvate is an intermediate in several metabolic pathways, but the majority is converted to acetyl-CoA and fed into the citric acid cycle. Although some more ATP is generated in the citric acid cycle, the most important product is NADH, which is made from NAD+ as the acetyl-CoA is oxidized. This oxidation releases carbon dioxide as a waste product. In anaerobic conditions, glycolysis produces lactate, through the enzyme lactate dehydrogenase re-oxidizing NADH to NAD+ for re-use in glycolysis. An alternative route for glucose breakdown is the pentose phosphate pathway, which reduces the coenzyme NADPH and produces pentose sugars such asribose, the sugar component of nucleic acids.
Fats are catabolised by hydrolysis to free fatty acids and glycerol. The glycerol enters glycolysis and the fatty acids are broken down bybeta oxidation to release acetyl-CoA, which then is fed into the citric acid cycle. Fatty acids release more energy upon oxidation than carbohydrates because carbohydrates contain more oxygen in their structures.
Amino acids are either used to synthesize proteins and other biomolecules, or oxidized to urea and carbon dioxide as a source of energy. The oxidation pathway starts with the removal of the amino group by a transaminase. The amino group is fed into the urea cycle, leaving a deaminated carbon skeleton in the form of a keto acid. Several of these keto acids are intermediates in the citric acid cycle, for example the deamination of glutamate forms α-ketoglutarate. The glucogenic amino acids can also be converted into glucose, through gluconeogenesis .
Much of Amino Acid Metabolism Involves Transamination
The amino acids are required for protein synthesis. Some must be supplied in the diet (the essential amino acids) since they cannot be synthesized in the body. The remainder are nonessential amino acids that are supplied in the diet but can be formed from metabolic intermediates by transamination, using the amino nitrogen from other amino acids. After deamination, amino nitrogen is excreted as urea, and the carbon skeletons that remain after transamination (1) are oxidized to CO2 via the citric acid cycle, (2) form glucose (gluconeogenesis), or (3) form ketone bodies.
Several amino acids are also the precursors of other compounds, eg, purines, pyrimidines, hormones such as epinephrine and thyroxine, and neurotransmitters.
Proteins are the main tissue builders in the body. They are part of every cell in the body. Proteins help in cell structure, functions, haemoglobin formation to carry oxygen, enzymes to carry out vital reactions and a myriad of other functions in the body. Proteins are also vital in supplying nitrogen for DNA and RNA genetic material and energy production.
Stages of catabolism
Catabolism can be broken down into 3 main stages.
Stage 1 – Stage of Digestion
The large organic molecules like proteins, lipids and polysaccharides are digested into their smaller components outside cells. This stage acts on starch, cellulose or proteins that cannot be directly absorbed by the cells and need to be broken into their smaller units before they can be used in cell metabolism.
Digestive enzymes include glycoside hydrolases that digest polysaccharides into monosaccharides or simple sugars.
The primary enzyme involved in protein digestion is pepsin which catalyzes the nonspecific hydrolysis of peptide bonds at an optimal pH of 2. In the lumen of the small intestine, the pancreas secretes zymogens of trypsin, chymotrypsin, elastase etc. These proteolytic enzymes break the proteins down into free amino acids as well as dipeptides and tripeptides. The free amino acids as well as the di and tripeptides are absorbed by the intestinal mucosa cells which subsequently are released into the blood stream where they are absorbed by other tissues.
The amino acids and sugars are then pumped into cells by specific active transport proteins.
Stage 2 – Release of energy
Once broken down these molecules are taken up by cells and converted to yet smaller molecules, usually acetyl coenzyme A (acetyl-CoA), which releases some energy.
Stage 3 - The acetyl group on the CoA is oxidised to water and carbon dioxide in the citric acid cycle and electron transport chain, releasing the energy that is stored by reducing the coenzyme nicotinamide adenine dinucleotide (NAD+) into NADH.
Carbohydrate breakdown
When complex carbohydrates are broken they form simple sugars or monosaccharides. This is taken up by the cells. Once inside these sugars undergo glycolysis, where sugars such as glucose and fructose are converted into pyruvate and some ATP is generated. Pyruvate is an intermediate in several metabolic pathways, but the majority is converted to acetyl-CoA and fed into the citric acid cycle or the Kreb’s cycle.
Within the citric acid cycle more ATP is generated by the monosaccharides. The most important product is NADH, which is made from NAD+ as the acetyl-CoA is oxidized. This oxidation releases carbon dioxide as a waste product.
When there is no oxygen, glycolysis produces lactate, through the enzyme lactate dehydrogenase, re-oxidizing NADH to NAD+ for re-use in glycolysis.
Glucose can also be broken down by pentose phosphate pathway, which reduces the coenzyme NADPH and produces pentose sugars such as ribose, the sugar component of nucleic acids.
Amino acid breakdown
Proteins are broken down into amino acids. Amino acids are either used to synthesize proteins and other biomolecules, or oxidized to urea and carbon dioxide as a source of energy.
In the process of oxidation, first the amino group is removed by a transaminase. The amino group is fed into the urea cycle, leaving a deaminated carbon skeleton in the form of a keto acid.
These keto acids enter the citric acid cycle. Glutamate, for example, forms α-ketoglutarate. Some of the amines may also be converted into glucose, through gluconeogenesis.
Some proteins are incredibly stable, others are very short lived. The short lived proteins usually play important metabolic roles. The short life times of these proteins allow the cell to rapidly adjust to changes in the metabolic state of the cell.
Lipid breakdown
Fats are catabolised by hydrolysis to free fatty acids and glycerol. The glycerol enters glycolysis and the fatty acids are broken down by beta oxidation to release acetyl-CoA. This acetyl co-A reaches the citric acid cycle next. Fatty acids release more energy upon oxidation than carbohydrates because carbohydrates contain more oxygen in their structures.
The chemical reactions of metabolism are organized into metabolic pathways. These allow the basic chemicals from nutrition to be transformed through a series of steps into another chemical, by a sequence of enzymes.
Enzymes are crucial to metabolism because they allow organisms to drive desirable reactions that require energy. These reactions also are coupled with those that release energy. As enzymes act as catalysts they allow these reactions to proceed quickly and efficiently. Enzymes also allow the regulation of metabolic pathways in response to changes in the cell's environment or signals from other cells.
Each metabolic pathway consists of a series of biochemical reactions that are connected by their intermediates: the products of one reaction are the substrates for subsequent reactions, and so on. Metabolic pathways are often considered to flow in one direction. Although all chemical reactions are technically reversible, conditions in the cell are often such that it is thermodynamically more favorable for flux to flow in one direction of a reaction. For example, one pathway may be responsible for the synthesis of a particular amino acid, but the breakdown of that amino acid may occur via a separate and distinct pathway. One example of an exception to this "rule" is the metabolism of glucose. Glycolysis results in the breakdown of glucose, but several reactions in the glycolysis pathway are reversible and participate in the re-synthesis of glucose (gluconeogenesis).
· Glycolysis was the first metabolic pathway discovered:
1. As glucose enters a cell, it is immediately phosphorylated by ATP to glucose 6-phosphate in the irreversible first step.
2. In times of excess lipid or protein energy sources, certain reactions in the glycolysis pathway may run in reverse in order to produce glucose 6-phosphate which is then used for storage as glycogen or starch.
· Metabolic pathways are often regulated by feedback inhibition.
· Some metabolic pathways flow in a 'cycle' wherein each component of the cycle is a substrate for the subsequent reaction in the cycle, such as in the Krebs Cycle (see below).
· Anabolic and catabolic pathways in eukaryotes often occur independently of each other, separated either physically by compartmentalization within organelles or separated biochemically by the requirement of different enzymes and co-factors.
Several distinct but linked metabolic pathways are used by cells to transfer the energy released by breakdown of fuel molecules into ATPand other small molecules used for energy (e.g. GTP, NADPH, FADH).
These pathways occur within all living organisms in some form:
1. Glycolysis
2. Aerobic respiration and/or Anaerobic respiration
3. Citric acid cycle / Krebs cycle (not in most obligate anaerobic organisms)
4. Oxidative phosphorylation (not in obligate anaerobic organisms)
Catabolism is characterized by convergence of three major routs toward a final common pathway.
Different proteins, fats and carbohydrates enter the same pathway – tricarboxylic acid cycle.
Anabolism can also be divided into stages, however the anabolic pathways are characterized by divergence.
Monosaccharide synthesis begin with CO2, oxaloacetate, pyruvate or lactate. Amino acids are synthesized from acetyl CoA, pyruvate or keto acids of Krebs cycle. .
Fatty acids are constructed from acetyl CoA.
On the next stage monosaccharides, amino acids and fatty acids are used for the synthesis of polysaccharides, proteins and fats.
Compartmentation of metabolic processes permits:
- separate pools of metabolites within a cell
- simultaneous operation of opposing metabolic paths
- high local concentrations of metabolites
Example: fatty acid synthesis enzymes (cytosol), fatty acid breakdown enzymes (mitochondria).
METABOLIC PATHWAYS MAY BE STUDIED AT DIFFERENT LEVELS
OF ORGANIZATION
In addition to studies in the whole organism, the location and integration of metabolic pathways is revealed by studies at several levels of organization. At the tissue and organ level, the nature of the substrates entering and metabolites leaving tissues and organs is defined. At the subcellular level, each cell organelle (eg, the mitochondrion) or compartment (eg, the cytosol) has specific roles that form part of a subcellular pattern of metabolic pathways.
At the Tissue and Organ Level, the Blood Circulation Integrates Metabolism
Amino acids resulting from the digestion of dietary protein and glucose resulting from the digestion of carbohydrate are absorbed and directed to the liver via the hepatic portal vein. The liver has the role of regulating the blood concentration of most water-soluble metabolites In the case of glucose, this is achieved by taking up glucose in excess of immediate requirements and converting it to glycogen.
Between meals, the liver acts to maintain the blood glucose concentration from glycogen (glycogenolysis) and, together with the kidney, by converting noncarbohydrate metabolites such as lactate, glycerol, and amino acids to glucose (gluconeogenesis). Maintenance of an adequate concentration of blood glucose is vital for those tissues in which it is the major fuel (the brain) or the only fuel (the erythrocytes).
The liver also synthesizes the major plasma proteins (eg, albumin) and deaminates amino acids that are in excess of requirements, forming urea, which is transported to the kidney and excreted. Skeletal muscle utilizes glucose as a fuel, forming both lactate and CO2. It stores glycogen as a fuel for its use in muscular contraction and synthesizes muscle protein from plasma amino acids. Muscle accounts for approximately 50% of body mass and consequently represents a considerable store of protein that can be drawn upon to supply amino acids for gluconeogenesis in starvation.
Lipids in the diet are mainly triacylglycerol and are hydrolyzed to monoacylglycerols and fatty acids in the gut, then reesterified in the intestinal mucosa. Here they are packaged with protein and secreted into the lymphatic system and thence into the
blood stream as chylomicrons, the largest of the plasma lipoproteins. Chylomicrons also contain other lipidsoluble nutrients, eg, vitamins. Unlike glucose and amino acids, chylomicron triacylglycerol is not taken up directly by the liver. It is first metabolized by tissues that have lipoprotein lipase, which hydrolyzes the triacylglycerol, releasing fatty acids that are incorporated into tissue lipids or oxidized as fuel. The other major source of long-chain fatty acid is synthesis (lipogenesis) from carbohydrate, mainly in adipose tissue and the liver. Adipose tissue triacylglycerol is the main fuel reserve of the body. On hydrolysis (lipolysis) free fatty acids are released into the circulation. These are taken up by most tissues (but not brain or erythrocytes) and esterified to acylglycerols or oxidized as a fuel. In the liver, triacylglycerol arising from lipogenesis, free fatty acids, and chylomicron remnants is secreted into the circulation as very low density lipoprotein (VLDL). This triacylglycerol undergoes a fate similar to that of chylomicrons. Partial oxidation of fatty acids in the liver leads to ketone body production Ketone bodies are transported to extrahepatictissues, where they act as a fuel source in starvation.
Pyruvate Dehydrogenase
Glycolysis enzymes are located in the cytosol of cells. Pyruvate enters the mitochondrion to be metabolized further.
Pyruvate dehydrogenase complex is a bridge between glycolysis and aerobic metabolism – citric acid cycle.
Flow diagram depicting the overall activity of the pyruvate dehydrogenase complex. During the oxidation of pyruvate to CO2 by pyruvate dehydrogenase the electrons flow from pyruvate to the lipoamide moiety of dihydrolipoyl transacetylase then to the FAD cofactor of dihydrolipoyl dehydrogenase and finally to reduction of NAD+ to NADH. The acetyl group is linked to coenzyme A (CoASH) in a high energy thioester bond. The acetyl-CoA then enters the TCA cycle for complete oxidation to CO2 and H2O.
Pyruvate freely diffuses through the outer membrane of mitochon-dria through the channels formed by transmembrane proteins porins.
Pyruvate Dehydrogenase catalyzes oxidative decarboxylation of pyruvate, to form acetyl-CoA. The overall reaction is shown below.
Pyruvate dehydrogenase complex is giant, with molecular mass ranging from 4 to 10 million daltons.
Pyruvate Dehydrogenase is a large complex containing many copies of each of three enzymes, E1, E2, and E3.
The inner core of the mammalian Pyruvate Dehydrogenase complex is an icosahedral structure consisting of 60 copies of E2.
At the periphery of the complex are:
· 30 copies of E1 (itself a tetramer with subunits a2b2) and
· 12 copies of E3 (a homodimer), plus 12 copies of an E3 binding protein that links E3 to E2.
Prosthetic groups are listed below
|
Enzyme |
Abbreviated |
Prosthetic Group |
|
Pyruvate Dehydrogenase |
E1 |
Thiamine pyrophosphate (TPP) |
|
Dihydrolipoyl Transacetylase |
E2 |
Lipoamide |
|
Dihydrolipoyl Dehydrogenase |
E3 |
FAD |
Thiamine pyrophosphate (TPP) is a derivative of thiamine (vitamin B1). Nutritional deficiency of thiamine leads to the disease beriberi. Beriberi affects especially the brain, because TPP is required for carbohydrate metabolism, and the brain depends on glucose metabolism for energy.
A proton readily dissociates from the C that is between N and S in the thiazole ring of TPP. The resulting carbanion (ylid) can attack the electron-deficient keto carbon of pyruvate.
Lipoamide includes a dithiol that undergoes oxidation and reduction.
The carboxyl group at the end of lipoic acid's hydrocarbon chain forms an amide bond to the side-chain amino group of a lysine residue of E2.
A long flexible arm, including hydrocarbon chains of lipoate and the lysine R-group, links the dithiol of each lipoamide to one of two lipoate-binding domains of each E2. Lipoate-binding domains are themselves part of a flexible strand of E2 that extends out from the core of the complex.
The long flexible attachment allows lipoamide functional groups to swing back and forth between E2 active sites in the core of the complex and active sites of E1 & E3 in the outer shell of the complex.
The E3 binding protein (that binds E3 to E2) also has attached lipoamide that can exchange reducing equivalents with lipoamide on E2.
FAD (Flavin Adenine Dinucleotide) is a derivative of the B-vitamin riboflavin (dimethylisoalloxazine-ribitol). The flavin ring system undergoes oxidation/reduction as shown below. Whereas NAD+ is a coenzyme that reversibly binds to enzymes, FAD is a prosthetic group, that is permanently part of the complex.
FAD accepts and donates 2 electrons with 2 protons (2 H):
FAD + 2 e- + 2 H+ �� FADH2
Organic arsenicals are potent inhibitors of lipoamide-containing enzymes such as Pyruvate Dehydrogenase. These highly toxic compounds react with "vicinal" dithiols such as the functional group of lipoamide as shown below.
In the overall reaction, the acetic acid generated is transferred to coenzyme A.
The final electron acceptor is NAD+.
The keto carbon of pyruvate reacts with the carbanion of TPP on E1 to yield an addition compound. The electron-pulling positively charged nitrogen of the thiazole ring promotes loss of CO2. What remains is hydroxyethyl-TPP.
The hydroxyethyl carbanion on TPP of E1 reacts with the disulfide of lipoamide on E2. What was the keto carbon of pyruvate is oxidized to a carboxylic acid, as the disulfide of lipoamide is reduced to a dithiol. The acetate formed by oxidation of the hydroxyethyl moiety is linked to one of the thiols of the reduced lipoamide as a thioester (~).
The acetate is transferred from the thiol of lipoamide to the thiol of coenzyme A, yielding acetyl CoA.
The reduced lipoamide swings over to the E3 active site. Dihydrolipoamide is reoxidized to the disulfide, as 2 e- + 2 H+ are transferred to a disulfide on E3 (disulfide interchange).
The dithiol on E3 is reoxidized as 2 e- + 2 H+ are transferred to FAD. The resulting FADH2 is reoxidized by electron transfer to NAD+, to yield NADH + H+.
Acetyl CoA, a product of the Pyruvate Dehydrogenase reaction, is a central compound in metabolism. The "high energy" thioester linkage makes it an excellent donor of the acetate moiety.
For example, acetyl CoA functions as:
· input to the Krebs Cycle, where the acetate moiety is further degraded to CO2.
· donor of acetate for synthesis of fatty acids, ketone bodies, and cholesterol.
The first enzyme of the complex is PDH itself which oxidatively decarboxylates pyruvate. During the course of the reaction the acetyl group derived from decarboxylation of pyruvate is bound to TPP. The next reaction of the complex is the transfer of the 2--carbon acetyl group from acetyl-TPP to lipoic acid, the covalently bound coenzyme of lipoyl transacetylase. The transfer of the acetyl group from acyl-lipoamide to CoA results in the formation of 2 sulfhydryl (SH) groups in lipoate requiring reoxidation to the disulfide (S-S) form to regenerate lipoate as a competent acyl acceptor. The enzyme dihydrolipoyl dehydrogenase, with FAD+ as a cofactor, catalyzes that oxidation reaction. The final activity of the PDH complex is the transfer of reducing equivalents from the FADH2 of dihydrolipoyl dehydrogenase to NAD+. The fate of the NADH is oxidation via mitochondrial electron transport, to produce 3 equivalents of ATP:
The net result of the reactions of the PDH complex are:
Pyruvate + CoA + NAD+ ------> CO2 + acetyl-CoA + NADH + H+
Regulation of the PDH Complex The reactions of the PDH complex serves to interconnect the metabolic pathways of glycolysis, gluconeogenesis and fatty acid synthesis to the TCA cycle. As a consequence, the activity of the PDH complex is highly regulated by a variety of allosteric effectors and by covalent modification. The importance of the PDH complex to the maintenance of homeostasis is evident from the fact that although diseases associated with deficiencies of the PDH complex have been observed, affected individuals often do not survive to maturity. Since the energy metabolism of highly aerobic tissues such as the brain is dependent on normal conversion of pyruvate to acetyl-CoA, aerobic tissues are most sensitive to deficiencies in components of the PDH complex. Most genetic diseases associated with PDH complex deficiency are due to mutations in PDH. The main pathologic result of such mutations is moderate to severe cerebral lactic acidosis and encephalopathies.
The main regulatory features of the PDH complex are diagrammed below.
|
|
|
Factors regulating the activity of pyruvate dehydrogenase, (PDH). PDH activity is regulated by its' state of phosphorylation, being most active in the dephosphorylated state. Phosphorylation of PDH is catalyzed by a specific PDH kinase. The activity of the kinase is enhanced when cellular energy charge is high which is reflected by an increase in the level of ATP, NADH and acetyl-CoA. Conversely, an increase in pyruvate strongly inhibits PDH kinase. Additional negative effectors of PDH kinase are ADP, NAD+ and CoASH, the levels of which increase when energy levels fall. The regulation of PDH phosphatase is not completely understood but it is known that Mg2+ and Ca2+ activate the enzyme. In adipose tissue insulin increases PDH activity and in cardiac muscle PDH activity is increased by catecholamines. |
Two products of the complex, NADH and acetyl-CoA, are negative allosteric effectors on PDH-a, the non-phosphorylated, active form of PDH. These effectors reduce the affinity of the enzyme for pyruvate, thus limiting the flow of carbon through the PDH complex. In addition, NADH and acetyl-CoA are powerful positive effectors on PDH kinase, the enzyme that inactivates PDH by converting it to the phosphorylated PDH-b form. Since NADH and acetyl-CoA accumulate when the cell energy charge is high, it is not surprising that high ATP levels also up-regulate PDH kinase activity, reinforcing down-regulation of PDH activity in energy-rich cells. Note, however, that pyruvate is a potent negative effector on PDH kinase, with the result that when pyruvate levels rise, PDH-a will be favored even with high levels of NADH and acetyl-CoA.
Concentrations of pyruvate which maintain PDH in the active form (PDH-a) are sufficiently high so that, in energy-rich cells, the allosterically down-regulated, high Km form of PDH is nonetheless capable of converting pyruvate to acetyl-CoA. With large amounts of pyruvate in cells having high energy charge and high NADH, pyruvate carbon will be directed to the 2 main storage forms of carbon (glycogen via gluconeogenesis and fat production via fatty acid synthesis) where acetyl-CoA is the principal carbon donor.
Although the regulation of
PDH-b phosphatase is not well understood, it is quite likely regulated to maximize
pyruvate oxidation under energy-poor conditions and to minimize PDH activity
under energy-rich conditions.
Regulation of Pyruvate Dehydrogenase complex.
Allosteric Regulation
Pyruvate dehydrogenase is a major regulatory point for entry of materials into the citric acid cycle.. The enzyme is regulated allosterically and by covalent modification.
E2 - inhibited by acetyl-CoA, activated by CoA-SH
E3 - inhibited by NADH, activated by NAD+.
ATP is an allosteric inhibitor of the complex, and AMP is an activator. The activity of this key reaction is coordinated with the energy charge, the [NAD+]/[NADH] ratio, and the ratio of acetylated to free coenzyme A.
Covalent Regulation
Part of the pyruvate dehydrogenase complex, pyruvate dehydrogenase kinase, phosphorylates three specific E1 serine residues, resulting in loss of activity of pyruvate dehydrogenase. NADH and acetyl-CoA both activate the kinase. The serines are dephosphorylated by a specific enzyme called pyruvate dehydrogenase phosphatase that hydrolyzes the phosphates from the E1 subunit of the pyruvate dehydgrogenase complex. This has the effect of activating the complex. The phosphatase is activated by Ca2+and Mg2+. Because ATP and ADP differ in their affinities for Mg2+, the concentration of free Mg2+ reflects the ATP/ADP ratio within the mitochondrion. Thus, pyruvate dehydrogenase responds to ATP levels by being turned off when ATP is abundant and further energy production is unneeded.
In mammalian tissues at rest, much less than half of the total pyruvate dehydrogenase is in the active, nonphosphorylated form. The complex can be turned on when low ATP levels signal a need to generate more ATP. The kinase protein is an integral part of the pyruvate dehydrogenase complex, whereas the phosphatase is but loosely bound.
At the Subcellular Level, Glycolysis Occurs in the Cytosol & the Citric Acid Cycle in the Mitochondria
Compartmentation of pathways in separate subcellular compartments or organelles permits integration and regulation of metabolism. Not all pathways are of equal importance in all cells. Depicts the subcellular compartmentation of metabolic pathways in a hepatic parenchymal cell.
The central role of the mitochondrion is immediately apparent, since it acts as the focus of carbohydrate, lipid, and amino acid metabolism. It contains the enzymes of the citric acid cycle, â-oxidation of fatty acids, and ketogenesis, as well as the respiratory chain and ATP synthase. Glycolysis, the pentose phosphate pathway, and fatty acid synthesis are all found in the cytosol. In gluconeogenesis, substrates such as lactate and pyruvate, which are formed in the cytosol, enter the mitochondrion to
yield oxaloacetate before formation of glucose. The membranes of the endoplasmic reticulum contain the enzyme system for acylglycerol synthesis, and the ribosomes are responsible for protein synthesis.
• The products of digestion provide the tissues with the building blocks for the biosynthesis of complex molecules and also with the fuel to power the living processes.
• Nearly all products of digestion of carbohydrate, fat, and protein are metabolized to a common metabolite, acetyl-CoA, before final oxidation to CO2 in the citric acid cycle.
• Acetyl-CoA is also used as the precursor for biosynthesis of long-chain fatty acids; steroids, including cholesterol; and ketone bodies.
• Glucose provides carbon skeletons for the glycerol moiety of fat and of several nonessential amino acids.
• Water-soluble products of digestion are transported directly to the liver via the hepatic portal vein. The liver regulates the blood concentrations of glucose and amino acids.
• Pathways are compartmentalized within the cell. Glycolysis, glycogenesis, glycogenolysis, the pentose phosphate pathway, and lipogenesis occur in the cytosol.
The mitochondrion contains the enzymes of the citric acid cycle, β-oxidation of fatty acids, and of oxidative phosphorylation. The endoplasmic reticulum also contains the enzymes for many other processes, including protein synthesis, glycerolipid
formation, and drug metabolism.
• Metabolic pathways are regulated by rapid mechanisms affecting the activity of existing enzymes, eg, allosteric and covalent modification (often in response
to hormone action); and slow mechanisms affecting the synthesis of enzymes.
Krebs Cycle
The Krebs cycle, also known as the tricarboxylic acid cycle (TCA), was first recognized in 1937 by the man for whom it is named, German biochemist Hans Adolph Krebs.
Sir Hans Adolf Krebs
Krebs was educated at the universities of
Göttingen, Freiburg,
Krebs is best known for his discovery of the Krebs
cycle (or tricarboxylic acid
cycle) in 1937. This is a continuation of the work of
Carl and Gerty Cori,
who had shown howcarbohydrates,
such as glycogen, are broken
down in the body to lactic acid;
Krebs completed the process by working out how the lactic acid is metabolized to carbon dioxideand
water. When he began this work little was known apart from the fact that the
process involved the consumption of oxygen, which could be increased, according
to AlbertSzent-Györgyi,
by the four-carbon compounds succinic acid, fumaric acid, malic acid,
and oxaloacetic acid.
Krebs himself showed in 1937 that the six-carbon citric acid is also
involved in the cycle.
By studying the process in pigeon breast muscle Krebs
was able to piece together the clues already collected into a coherent scheme. The
three-carbon lactic acid is first broken down to a two-carbon molecule
unfamiliar to Krebs; it was in fact later identified by Fritz Lipmann as coenzyme A. This
then combines with the four-carbon oxaloacetic acid to form the six-carbon citric acid. The
citric acid then undergoes a cycle of reactions to be converted to oxaloacetic
acid once more. During this cycle two molecules of carbondioxide are given up
and hydrogen atoms are released; the hydrogen is then oxidized in the electron
transport chain with the production of energy. Much of the detail of this aspect
of the cycle was later filled in by Lipmann, with whom Krebs shared the 1953
Nobel Prize for physiology or medicine.
Krebs fully appreciated the significance of the cycle, pointing out the important fact that it is the common terminal pathway for the chemical breakdown of all foodstuffs.
In 1932, with K. Henselheit, Krebs was responsible for the introduction of another cycle. This was the urea cycle, whereby amino acids (the constituents of proteins) eliminate their nitrogen in the form of urea, which is excreted in urine. This left the remainder of the amino acid to give up its potential energy and participate in a variety of metabolic pathways.
Hans A. Krebs, the son of Georg Krebs, an
otolaryngologist, was born in Hildesheim,
In 1935 Krebs went to the
The Ornithine Cycle
To keep organs and tissues alive for biochemical tests, they had been perfused with physiological salines as a substitute for blood. The results were often unsatisfactory. Early in his career Krebs devised the tissue-slice technique. The organ, rapidly removed after the death of the test animal, was cut into thin slices and kept in fresh saline forbiochemical testing. He used this technique in his study of the synthesis of urea by the liver.
It was known that urea is produced in a liver undergoing autolysis, and in 1904 it was shown that the autolysis produces the amino acid arginine, which is acted on catalytically by the enzyme arginase to produce urea. In 1932 Krebs found that, when an amino acid is added to liver, ammonia is liberated and is converted approximately quantitatively into urea. All the amino acids tested gave this result except two. When ornithine was added, the urea production was 10 times the expected amount, and arginine also gave an excess yield of urea. He therefore suggested that ornithine reacted with added ammonia and carbon dioxide to form arginine. Under the action of arginase, the arginine was broken down to urea and ornithine. If ammonia was omitted, there was no appreciable formation of urea. Further, ornithine was not observed to disappear while, with added ammonia, the synthesis of urea was in progress. Krebs therefore concluded that the ornithine acted as a catalyst. Many other substances were tested, but the only one that acted like ornithine was citrulline, and he suggested that citrulline formed a stage midway between ornithine and arginine. His ornithine cycle is still regarded as a sound explanation of the synthesis of urea in the body.
The Citric Acid Cycle
Krebs then turned to the intermediary oxidation of carbohydrates. In 1935 Albert von Szent-Györgyi elucidated the sequence of oxidations of the C4-dicarboxylic acids as follows:
succinic acid→fumaric acid→malic acid→maoxaloacetic acid
He also showed that these reactions were at least in part catalytic. This was later proved, but the manner of action remained unknown. In 1936 C. Martius and F. Knoop showed that in biological material citrate yields alphaketoglutarate on oxidation. They further suggested that the intermediate products were cis -aconitic acid, isocitric acid, andoxalosuccinic acid. It was already known that alpha-ketoglutarate forms succinate. In 1937, when Krebs started his work, the following sequence of reactions was therefore known:
citric acid→cis -aconitic acid→iso-citric acid→oxalosuccinic acid→alpha-ketoglutamic acid→succinic acid→fumaric acid→malic acid→oxaloacetic acid
Krebs and W. A. Johnson found that citrate was not only rapidly broken down in muscle but was also readily formed provided that oxaloacetate was added. The assumption was that some of the oxaloacetate was broken down to pyruvate or acetate and that the formation of citrate was due to a combination of the remaining oxaloacetate with pyruvate or acetate. But pyruvate or acetate could be derived from carbohydrate. In 1937 Krebs conceived the whole process as a cycle in which an undefined derivative of pyruvate, resulting from the breakdown of carbohydrate, condensed with oxaloacetate to form citric acid. The citric acid then passed through the changes noted above untiloxaloacetic acid was regenerated, and the cycle was repeated. The full cycle is therefore as follows:
citric acid→cis -aconitic acid→iso-citric acid→oxalosuccinic acid→alpha-ketoglutamic acid→succinic acid→fumaric acid→malic acid→oxaloacetic acid+pyruvic acid→citric acid
Since Krebs originally described this cycle, he and others did further work on it. In 1950 Fritz Lipmann showed that the derivative of pyruvic acid that combines with oxaloacetate to form citrate is acetyl-coenzyme A and that this coenzyme is also active at two other points in the cycle. It was shown that acetyl-coenzyme A, in addition to its formation from carbohydrate, is also formed from fatty acids and many amino acids. The Krebs cycle is therefore a most important concept of biochemistry. Krebs shared with Lipmann the Nobel Prize in Physiology or Medicine in 1953.
Among Krebs's other important contributions to biochemistry were his studies of the synthesis of glutamine in brain tissue under the influence of the enzyme glutaminase(1935), the passage of ions across cell membranes (1950), and the effect of primitive intrinsic regulating mechanisms in controlling the metabolism of metazoan cells (1957).
Later Life
In 1967 Krebs, having reached
Krebs received many honors in addition to his Nobel Prize. In 1947 he was elected a Fellow of the Royal Society, and he was awarded its Royal (1954) and Copley (1961) Medals. He delivered its Croonian Lecture in 1963. He was a member of many foreign scientific societies, and he held honorary doctorates from 14 universities. He received the Gold Medal of the Royal Society of Medicine in 1965, and he was knighted in 1958.
Read more: http://www.answers.com/topic/hans-adolf-krebs#ixzz2QQAAV519
http://www.youtube.com/watch?v=A1DjTM1qnPM&feature=related
The Krebs cycle refers to a complex series of chemical reactions that produce carbon dioxide and Adenosine triphosphate (ATP), a compound rich in energy. The cycleoccurs by essentially linking two carbon coenzyme with carbon compounds; the created compound then goes through a series of changes that produce energy. This cycle occurs in all cells that utilize oxygen as part of their respiration process; this includes those cells of creatures from the higher animal kingdom such as humans. Carbon dioxide is important for various reasons, the main one being that it stimulates breathing, while ATP provides cells with the energy required for the synthesis of proteins from amino acids and the replication of deoxyribonucleic acid (DNA); both are vital for energy supply and for life to continue. In short, the Krebs cycle constitutes the discovery of the major source of energy in all living organisms.
Functions
Within the Krebs cycle, energy in the form of ATP is usually derived from the breakdown of glucose, although fats and proteins can also be utilized as energy sources. Since glucose can pass through cell membranes, it transports energy from one part of the body to another. The Krebs cycle affects all types of life and is, as such, the metabolic pathway within the cells. This pathway chemically converts carbohydrates, fats, and proteins into carbon dioxide, and converts water into serviceable energy.
The Krebs cycle is the second stage of aerobic respiration, the first being glycolysis and last being the electron transport chain; the cycle is a series of stages that every living cell must undergo in order to produce energy. The enzymes that cause each step of the process to occur are all located in the cell's "power plant"; in animals, this power plant is the mitochondria; in plants, it is the chloroplasts; and in microorganisms, it can be found in the cell membrane. The Krebs cycle is also known as the citric acid cycle, because citric acid is the very first product generated by this sequence of chemical conversions, and it is also regenerated at the end of the cycle.
The pyruvate molecules produced during glycolysis contains a lot of energy in the bonds between their molecules. In order to use that energy, the cell must convert it into the form of ATP. To do so, pyruvate molecules are processed through the Kreb Cycle, also known as the citric acid cycle.
http://www.youtube.com/watch?v=7gR4s8ool1Y
(Kerbs Cycle as a drawing)
1. Prior to entering the Krebs Cycle, pyruvate must be converted into acetyl CoA. This is achieved by removing a CO2 molecule from pyruvate and then removing an electron to reduce an NAD+ into NADH. An enzyme called coenzyme A is combined with the remaini ow:
2. Citrate is formed when the acetyl group from acetyl CoA combines with oxaloacetate from the previous Krebs cycle.
3. Citrate is converted into its isomer isocitrate.
4. Isocitrate is oxidized to form the 5-carbon α-ketoglutarate. This step releases one molecule of CO2 and reduces NAD+ to NADH2+.
5. The α-ketoglutarate is oxidized to succinyl CoA, yielding CO2 and NADH2+.
The a-Ketoglutarate Dehydrogenase Complex is
Similar to pyruvate dehydrogenase complex
Same coenzymes, identical mechanisms
E1 - a-ketoglutarate dehydrogenase (with TPP)
E2 – dihydrolipoyl succinyltransferase (with flexible lipoamide prosthetic group)
E3 - dihydrolipoyl dehydrogenase (with FAD)
6. Succinyl CoA releases coenzyme A and phosphorylates ADP into ATP.
In the succinyl CoA synthetase reaction, the thioester bond between HS-CoA and the succinyl group is hydrolyzed.
Since it is a rich in energy bond, the energy released is enough for synthesizing GTP from GDP + (P).
This GTP is equivalent, from the energetic point of view, to ATP. In fact, GTP can transfer the (P) group to ADP to form ATP:
GTP + ADP ————–à GDP + ATP
Since ATP can be produced from this reaction, without participation of the respiratory chain, this process is called Substrate Level Phosphorylation (SLP) in contrast to the Oxidative Phosphorylation (ATP synthesis using the energy released in the Electron Transport Chain).
A few other reactions in metabolism are also coupled with ATP synthesis without participation of the respiratory chain. They are considered also SLP reactions.
7. Succinate is oxidized to fumarate, converting FAD to FADH2.
The Succinate Dehydrogenase Complex of several polypeptides, an FAD prosthetic group and iron-sulfur clusters, embedded in the inner mitochondrial membrane. Electrons are transferred from succinate to FAD and then to ubiquinone (Q) in electron transport chain. Dehydrogenation is stereospecific; only the trans isomer is formed
8. Fumarate is hydrolized to form malate.
9. Malate is oxidized to oxaloacetate, reducing NAD+ to NADH2+.
We are now back at the beginning of the Krebs Cycle. Because glycolysis produces two pyruvate molecules from one glucose, each glucose is processes through the kreb cycle twice. For each molecule of glucose, six NADH2+, two FADH2, and two ATP.

{kind=link}
Overview
The sum of all reactions in the citric acid cycle is:
Acetyl-CoA + 3 NAD+ + FAD + GDP + Pi + 2 H2O → CoA-SH + 3 NADH + 3 H+ + FADH2 + GTP + 2 CO2
(the above reaction is equilibrated if Pi represents the H2PO4- ion, GDP the GDP2- ion and GTP the GTP3- ion).
Two carbons are oxidized to CO2, and the energy from these reactions is stored in GTP, NADH and FADH2. NADH and FADH2 are coenzymes (molecules that enable or enhance enzymes) that store energy and are utilized in oxidative phosphorylation.
A simplified view of the process
· Electrons are also transferred to the electron acceptor FAD, forming FADH2.
· At the end of all cycles, the products are two GTP, six NADH, two FADH2, four CO2.
http://www.youtube.com/watch?v=hw5nWB0xN0Y&feature=related
INVESTIGATION OF BIOLOGICAL OXIDATION, OXIDATIVE PHOSPHORYLATION AND ATP SYNTHESIS. INHIBITORS AND UNCOUPLERS OF OXIDATIVE PHOSPHORYLATION.
Bioenergetic
Bioenergetics is the part of biochemistry concerned with the energy involved in making and breaking of chemical bonds in the molecules found in biological organisms.
Growth, development and metabolism are some of the central phenomena in the study of biological organisms. The role of energy is fundamental to such biological processes. The ability to harness energy from a variety of metabolic pathways is a property of all living organisms. Life is dependent on energy transformations; living organisms survive because of exchange of energy within and without.
In a living organism, chemical bonds are broken and made as part of the exchange and transformation of energy. Energy is available for work (such as mechanical work) or for other processes (such as chemical synthesis and anabolic processes in growth), when weak bonds are broken and stronger bonds are made. The production of stronger bonds allows release of usable energy.
Living organisms obtain energy from organic and inorganic materials. For example, lithotrophs can oxidize minerals such as nitrates or forms of sulfur, such as elemental sulfur, sulfites, and hydrogen sulfide to produce ATP. In photosynthesis, autotrophs can produce ATP using light energy. Heterotrophs must consume organic compounds. These are mostly carbohydrates, fats, and proteins. The amount of energy actually obtained by the organism is lower than the amount present in the food; there are losses in digestion, metabolism, and thermogenesis.
The materials are generally combined with oxygen to release energy, although some can also be oxidized anaerobically by various organisms. The bonds holding the molecules of nutrients together and the bonds holding molecules of free oxygen together are all relatively weak compared with the chemical bonds holding carbon dioxide and water together. The utilization of these materials is a form of slow combustion. That is why the energy content of food can be estimated with a bomb calorimeter. The materials are oxidized slowly enough that the organisms do not actually produce fire. The oxidation releases energy because stronger bonds have been formed. This net energy may evolve as heat, or some of which may be used by the organism for other purposes, such as breaking other bonds to do chemistry.
Living organisms produce ATP from energy sources via oxidative phosphorylation. The terminal phosphate bonds of ATP are relatively weak compared with the stronger bonds formed when ATP is broken down to adenosine monophosphate and phosphate, dissolved in water. Here it is the energy of hydration that results in energy release. This hydrolysis of ATP is used as a battery to store energy in cells, for intermediate metabolism. Utilization of chemical energy from such molecular bond rearrangement powers biological processes in every biological organism.
Exergonic and endergonic processes. Mechanism of energy releasing and storage in the organism.
Exergonic refers to chemical reactions that proceed spontaneously from reactants to products with the release of energy. Endergonic reactions require energy input to proceed. Although the terms are often used rather loosely, they are precisely defined thermodynamic concepts based on changes in an entity called Gibbs free energy (G) accompanying reactions. Reactions in which -G decreases are exergonic, and those in which -G increases are endergonic. Exergonic reactions often involve the breakdown of organic compounds found in food, whereas endergonic reactions frequently entail synthesis of complicated molecules.
Biological metabolism contains many examples of both types, and living organisms have developed elaborate techniques for coupling the two.
Exergonic reactions release free energy while endergonic reactions consume free energy
Although a negative -G indicates that energy must be added to the system before a reaction will occur, it tells us nothing about the rate at which it will progress. As is often the case, it may go very slowly if substantial activation energy is required to start the reaction. Living organisms have found a way around this problem by forming protein catalysts, called enzymes, that effectively reduce the amount of activation energy needed, and allow the reaction to proceed at a satisfactory rate. Enzymes do not affect the free energy of the reaction, and will not enable reactions to proceed that are not energetically feasible.
By coupling exergonic and endergonic reactions, organisms are able to use the available energy in food they consume to construct complex proteins, lipids, nucleic acids and carbohydrates needed for their growth and development. A well-known example involves coupling the formation of energy-rich adenosine triphosphate (ATP) from adenosine diphosphate (ADP) and phosphate (an endergonic reaction), with the transfer of hydrogen, removed from organic food materials, to oxygen (an exergonic reaction). The process is called oxidative phosphorylation. Energy stored in ATP may be used subsequently when the exergonic conversion of ATP back to ADP and phosphate is coupled with the endergonic synthesis of a needed cellular component.
What is macroergic bond? Examples of high energy compounds.
A bond in chemical compounds, which acts as an energy accumulator. Macroergic bond is present in some phosphorus-containing compounds in living organisms, e. g.* adenosinetriphosphate. Macroergic bonds are formed as a result of complex biochemical processes and break up when energy is released. The processes are reversible and can be repeated.
High-energy phosphate can mean one of two things:
· The phosphate-phosphate bonds formed when compounds such as adenosine diphosphate and adenosine triphosphate are created.
· The compounds that contain these bonds, which include the nucleoside diphosphates and nucleoside triphosphates, and the high-energy storage compounds of the muscle, the phosphagens. When people speak of a high-energy phosphate pool, they speak of the total concentration of these compounds with these high-energy bonds.
·
High-energy phosphate bonds are pyrophosphate bonds, acid anhydride linkages formed by taking phosphoric acid derivatives and dehydrating them. As a consequence, the hydrolysis of these bonds is exergonic under physiological conditions, releasing energy.
|
Energy released by high energy phosphate reactions |
|
|
Reaction |
ΔG [kJ/mol] |
|
ATP + H2O → ADP + Pi |
-30.5 |
|
ADP + H2O → AMP + Pi |
-30.5 |
|
ATP + H2O → AMP + PPi |
-40.6 |
|
PPi + H2O → 2 Pi |
-31.8 |
|
AMP + H2O → A + Pi |
-12.6 |
Except for PPi → 2 Pi, these reactions are, in general, not allowed to go uncontrolled in the human cell but are instead coupled to other processes needing energy to drive them to completion. Thus, high-energy phosphate reactions can:
· provide energy to cellular processes, allowing them to run;
· couple processes to a particular nucleoside, allowing for regulatory control of the process;
· drive the reaction to the right, by taking a reversible process and making it irreversible.
The one exception is of value because it allows a single hydrolysis, ATP + 2H2O → AMP + PPi, to effectively supply the energy of hydrolysis of two high-energy bonds, with the hydrolysis of PPi being allowed to go to completion in a separate reaction. The AMP is regenerated to ATP in two steps, with the equilibrium reaction ATP + AMP ↔ 2ADP, followed by regeneration of ATP by the usual means, oxidative phosphorylation or other energy-producing pathways such as glycolysis.
Often, high-energy phosphate bonds are denoted by the character '~'. In this "squiggle" notation, ATP becomes A-P~P~P. The squiggle notation was invented by Fritz Albert Lipmann, who first proposed ATP as the main energy transfer molecule of the cell, in 1941. It emphasizes the special nature of these bonds.
Stryer states:
ATP is often called a high energy compound and its phosphoanhydride bonds are referred to as high-energy bonds. There is nothing special about the bonds themselves. They are high-energy bonds in the sense that free energy is released when they are hydrolyzed, for the reasons given above.
Lipmann’s term “high-energy bond” and his symbol ~P (squiggle P) for a compound having a high phosphate group transfer potential are vivid, concise, and useful notations. In fact Lipmann’s squiggle did much to stimulate interest in bioenergetics.
The term 'high energy' with respect to these bonds can be misleading because the negative free energy change is not due directly to the breaking of the bonds themselves. The breaking of these bonds, as with the breaking of most bonds, is an endergonic step (i.e., it absorbs energy, not releases it). The negative free energy change comes instead from the fact that the bonds formed after hydrolysis-or the phosphorylation of a residue by ATP-are lower in energy than the bonds present before hydrolysis (this includes all of the bonds involved in the reaction, not just the phosphate bonds themselves). This effect is due to a number of factors including increased resonance stabilization and solvation of the products relative to the reactants.
Besides the adenosine nucleotide phosphates, uracil, cytosine and guanine phosphates occur, too:
UMP, UDP, UTP, CMP, CDP, CTP, GMP, GDP, GTP.
The triphosphate nucleosides of these compounds and those of ATP are components of RNA. They are integrated into the polymer by splitting off pyrophosphate ( = PP). The corresponding desoxyribose derivatives (dATP, dGTP, dCTP....) are necessary for DNA synthesis, where dTTP is used instead of dUTP. The terminal phosphate residues of all nucleoside di- and triphosphates are equally rich in energy. The energy set free by their hydrolysis is used for biosyntheses. They share the work equally: UTP is needed for the synthesis of polysaccharides, CTP for that of lipids and GTP for the synthesis of proteins and other molecules. These specificities are the results of the different selectivities of the enzymes, that control each of these metabolic pathways.
ATP formation
The Function of ATP
The ATP is used for many cell functions including transport work moving substances across cell membranes. It is also used for mechanical work, supplying the energy needed for muscle contraction. It supplies energy not only to heart muscle (for blood circulation) and skeletal muscle (such as for gross body movement), but also to the chromosomes and flagella to enable them to carry out their many functions. A major role of ATP is in chemical work, supplying the needed energy to synthesize the multi-thousands of types of macromolecules that the cell needs to exist.
ATP is also used as an on-off switch both to control chemical reactions and to send messages. The shape of the protein chains that produce the building blocks and other structures used in life is mostly determined by weak chemical bonds that are easily broken and remade. These chains can shorten, lengthen, and change shape in response to the input or withdrawal of energy. The changes in the chains alter the shape of the protein and can also alter its function or cause it to become either active or inactive.
The ATP molecule can bond to one part of a protein molecule, causing another part of the same molecule to slide or move slightly which causes it to change its conformation, inactivating the molecule. Subsequent removal of ATP causes the protein to return to its original shape, and thus it is again functional. The cycle can be repeated until the molecule is recycled, effectively serving as an on and off switch. Both adding a phosphorus (phosphorylation) and removing a phosphorus from a protein (dephosphorylation) can serve as either an on or an off switch.
How is ATP Produced?
ATP is manufactured as a result of several cell processes including fermentation, respiration and photosynthesis. Most commonly the cells use ADP as a precursor molecule and then add a phosphorus to it. In eukaryotes this can occur either in the soluble portion of the cytoplasm (cytosol) or in special energy-producing structures called mitochondria. Charging ADP to form ATP in the mitochondria is called chemiosmotic phosphorylation. This process occurs in specially constructed chambers located in the mitochondrion’s inner membranes.
ATP-synthase converts ADP into ATP, a process called charging.
Adenosine triphosphate (ATP) is an organic molecule which stores energy used to carry out life processes. ATP is made of an adenine nucleoside, ribose sugar, and three phosphate groups. The high energy bonds between phosphate groups are broken when hydrolyzed, thus releasing energy in the system. Either one or two phosphate groups can break off, releasing Gibb's free energy, which can then be used to drive other reactions.
The molecular structure of ATP which is formed from a adenine nucleoside, ribose sugar, and three phosphate groups
ATP can be formed from bonding either adenosine monophosphate (AMP) and two inorganic phosphate groups (PPi) together or by bonding adenosine diphosphate (ADP) and one inorganic phosphate group (Pi) together. Energy is required to bond the adenosine to the phosphate groups, making it an endergonic reaction. The energy used to bond the two molecules together is then stored within covalent bonds between phosphate groups in ATP. ATP can be formed through two different endergonic processes, either through substrate-level phosphorylation or chemiosmosis.
ATP is needed
- as a source of energy for biochemical syntheses
- for transport processes (active transport) and
- for mechanical work like movements (ciliar movements, plasma currents etc.)
How the Hydrolysis of ATP Performs Work
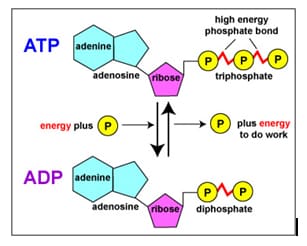
•The bonds between the phosphate groups of ATP’s tail can be broken by hydrolysis
•Energy is released from ATP when the terminal phosphate bond is broken
•This release of energy comes from the chemical change to a state of lower free energy, not from the phosphate bonds themselves
•The three types of cellular work (mechanical, transport, and chemical) are powered by the hydrolysis of ATP
•In the cell, the energy from the exergonic reaction of ATP hydrolysis can be used to drive an endergonic reaction
•Overall, the coupled reactions are exergonic
ATP + H2O → ADP + Pi
Releases -30.5 kJ/mol= ΔG˚ (when one phosphate group breaks off)
ATP + H2O → AMP + PPi
Releases -45.6 kJ/mol= ΔG˚ (when two phosphate groups break off)
Hydrolysis of ATP
The Regeneration of ATP
•ATP is a renewable resource that is regenerated by addition of a phosphate group to adenosine diphosphate (ADP)
•The energy to phosphorylate ADP comes from catabolic reactions in the cell
•The ATP cycle is a revolving door through which energy passes during its transfer from catabolic to anabolic pathways
http://www.youtube.com/watch?v=_PgjsfY71AM&feature=related
http://www.youtube.com/watch?v=YndC0gS3t6M&feature=related
Substrate-level and oxidative phosphorylation
Substrate-level phosphorylation is a type of metabolic reaction that results in the formation of adenosine triphosphate (ATP) or guanosine triphosphate (GTP) by the direct transfer and donation of a phosphoryl (PO3) group to adenosine diphosphate (ADP) or guanosine diphosphate (GDP) from a phosphorylated reactive intermediate. Note that the phosphate group does not have to come directly from the substrate. By convention, the phosphoryl group that is transferred is referred to as a phosphate group.
An alternative way to create ATP is through oxidative phosphorylation, which takes place during the process of cellular respiration, in addition to the substrate-level phosphorylation that occurs during glycolysis and the Krebs cycle. During oxidative phosphorylation, NADH is oxidized to NAD+, yielding 2.5 ATPs, and FADH2 yields 1.5 ATPs when it is oxidized. Oxidative phosphorylation uses an electrochemical or chemiosmotic gradient of protons (H+) across the inner mitochondrial membrane to generate ATP from ADP, which is a key difference from substrate-level phosphorylation.
Unlike oxidative phosphorylation, oxidation and phosphorylation are not coupled in the process of substrate-level phosphorylation, although both types of phosphorylation result in the formation of ATP and reactive intermediates are most often gained in course of oxidation processes in catabolism. However, usually most of the ATP is generated by oxidative phosphorylation in aerobic or anaerobic respiration. Substrate-level phosphorylation serves as fast source of ATP independent of external electron acceptors and respiration. This is the case for example in human erythrocytes, which have no mitochondria, and in the muscle during oxygen depression.

oxidative phosphorylation
The main part of substrate-level phosphorylation occurs in the cytoplasm of cells as part of glycolysis and in mitochondria as part of the Krebs Cycle under both aerobic and anaerobic conditions. In the pay-off phase of glycolysis, two ATP are produced by substrate-level phosphorylation: two and only two 1,3-bisphosphoglycerate are converted to 3-phosphoglycerate by transferring a phosphate group to ADP by a kinase; two phosphoenolpyruvate are converted to pyruvate by the transfer of their phosphate groups to ADP by another kinase. The first reaction occurs after the generation of 1,3-bisphosphoglycerate from 3-phosphoglyceraldehyde and an organic phosphate via a dehydrogenase.
ATP is generated in a following separate step (key difference from oxidative phosphorylation) by transfer of the high-energy phosphate on 1,3-bisphosphoglycerate to ADP via the enzyme phosphoglycerate kinase, generating 3-phosphoglycerate. As ATP is formed of a former inorganic phosphate group, this step leads to the energy yield of glycolysis. The second substrate-level phosphorylation occurs later by means of the reaction of phosphenolpyruvate (PEP) to pyruvate via the pyruvate kinase. This reaction regenerates the ATP that has been used in the preparatory phase of glycolysis to activate glucose to glucose-6-phosphate and fructose-6-phosphate to fructose-1,6-bisphosphate, respectively.
ATP can be generated by substrate-level phosphorylation in the mitochondrial matrix, a pathway that is independent from the protonmotive force, pmf. In the mitochondrial matrix there are two reactions capable of substrate-level phosphorylation: the mitochondrial phosphoenolpyruvate carboxykinase (PEPCK), and the succinate-CoA ligase (SUCL or succinate thiokinase or succinyl-CoA synthetase). Mitochondrial PEPCK is thought to participate in the transfer of the phosphorylation potential from the matrix to cytosol and vice versa. The enzyme is a heterodimer, being composed of an invariant alpha subunit encoded by SUCLG1, and a substrate-specific beta subunit, encoded by either SUCLA2 or SUCLG2. This dimer combination results in either an ADP-forming succinate-CoA ligase (A-SUCL, EC 6.2.1.5) or a GDP-forming succinate-CoA ligase (G-SUCL, EC 6.2.1.4). The ADP-forming succinate-CoA ligase is potentially the only matrix enzyme generating ATP in the absence of a pmf, capable of maintaining matrix ATP levels under energy-limited conditions, such as transient hypoxia.
Another form of substrate-level phosphorylation is also seen in working skeletal muscles and the brain. Phosphocreatine is stored as a readily available high-energy phosphate supply, and the enzyme creatine phosphokinase transfers a phosphate from phosphocreatine to ADP to produce ATP. Then the ATP releases giving chemical energy.
Apart from this substrate-level phosphorylation can also be observed in fermentation, for example, heterolactic fermentation, butyric acid fermentation, and propanoic acid fermentation.
The modern views on the biological oxidation
Biological oxidation is that oxidation which occurs in biological systems to produce energy.
Oxidation can occur by:
1-Addition of oxygen (less common)
2-Removal of hydrogen (common)
3-Removal of electrons (most common)
Electrons are not stable in the free state, so their removal form a substance (oxidation) must be accompanied by their acceptance by another substance (reduction) hence the reaction is called oxidation-reduction reaction or redox reaction and the involved enzymes are called oxido-reductases
Redoxpotential
It is the affinity of a substance to accept electrons i.e. it is the potential for a substance to become reduced. Hydrogen has the lowest redoxpotential (-0.42 volt), while oxygen has the highest redoxpotential (+0.82 volt). The redoxpotentials of all other substances lie between that of hydrogen and oxygen.
Electrons are transferred from substances with low redoxpotential to substances with higher redoxpotential.This transfer of electrons is an energy yielding process and the amount of energy liberated depends on the redoxpotential difference between the electron donor and acceptor.
Oxido-reductases
These enzymes catalyze oxidation-reduction reactions.

They are classified into five groups:
1-oxidases. 2-aerobic dehydrogenises.
3-anaerobic dehydrogenises.
4-hydroperoxidasesand
5-oxygenases.
1. Oxidases
An oxidase is any enzyme that catalyzes an oxidation-reduction reaction involving molecular oxygen (O2) as the electron acceptor. In these reactions, oxygen is reduced to water (H2O) or hydrogen peroxide (H2O2). The oxidases are a subclass of the oxidoreductases.
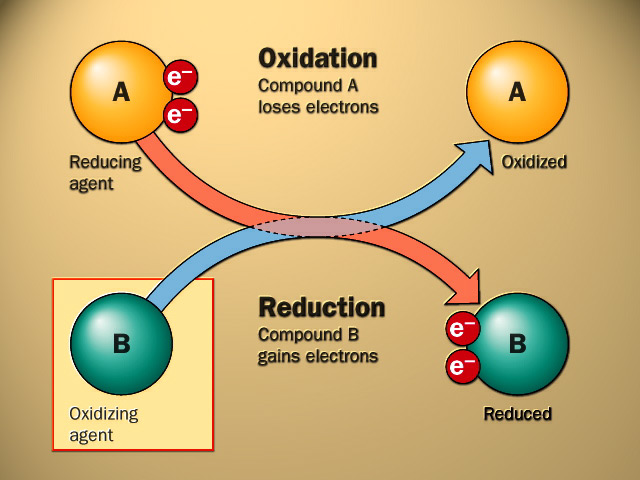
OXIDATION-REDUCTION OR REDOX REACTIONS
2. Aerobic Dehydrogenases(FlavoproteinLinked Oxidases).
The coenzyme of aerobic dehydrogenasesmay be:
•FMN (Flavinadenine mononucleotide) as in L-amino acid oxidase.
•FAD (Flavinadenine dinucleotide) as in D-amino acid oxidase, xanthineoxidase, aldehydedehydrogenaseand glucose oxidase.
3. Anaerobic Dehydrogenases
Anaerobic dehydrogenasesare further classified according to their coenzymes into:
•NAD+linked anaerobic dehydrogenasese.g.
a)Cytoplasmicglycerol-3-phosphate dehydrogenase
b)Isocitratedehydrogenase.
c)Malatedehydrogenase.
d)β-HydroxyacylCoAdehydrogenase.
e)β-Hydroxybutyrate dehydrogenase.
•NADP linked anaerobic dehydrogenasese.g.
a)Glucose-6-phosphate dehydrogenase.
b)Malicenzyme.c)Cytoplasmicisocitratedehydrogenase
•FAD linked anaerobic dehydrogenasese.g.
a)Succinatedehydrogenase.
b)Mitochondrial glycerol-3-phosphate dehydrogenase.
c)Acy1 CoAdehydrogenase.
4. Hydroperoxidases
These enzymes use hydrogen peroxide (H2O2) as substrate changing it into water to get rid of its harmful effects.
They are further classified into peroxidasesand catalases.
•Peroxidases: These enzymes need a reduced substrate as hydrogen donorperoxidase
H2O2 + XH2 (reduced substrate) ----------→ X(oxidized substrate)+ 2H2O Example:
-Glutathione peroxidasegets rid of H2O2from red cells to protect them from haemolysis
Glutathione Peroxidase
H2O2 + 2 G-S H -----------→2H2O + G-S-S-G
•Catalases:These enzymes act on 2 molecules of hydrogen peroxide; one molecule is hydrogen donor & the other molecule is hydrogenaccepetor.
2H2O2 + catalase---------→ 2H2O + O2
Hydrogen peroxide is continuously produced by the action of aerobic dehydrogenasesand some oxidases. It is also produced by action of superoxide dismutase on superoxide (O•2). It is removed by the action of peroxidasesand catalasesto protect cells against its harmful effects.
5. Oxygenases
These enzymes catalyze direct incorporation (addition) of oxygen into substrate.
They are further classified into dioxygenasesand monooxygenases.
A. Dioxygenases(true oxgenases)
These enzymes catalyze direct incorporation of two atoms of oxygen molecule into substrate e.g. tryptophan pyrrolase, homnogentisicacid dioxygenase, carotenaseand β-hydroxyanthranilicacid dioxygenase.
Dioxygenase
A + O2 → AO2
B. Mono-oxygenases(pseudo-oxygenases; hydroxylases; mixed: function oxygenases)
AH + O2+ XH2 ------→A-OH + H2O + X
Fuctionsof cytochromeP450
Functions of microsomalcytochromeP450
1-It is important for detoxicationof xenobioticsby hydroxylation. e.g. insecticides,carcinogens,mutagensand drugs.
2-It is also important for metabolism of some drugs by hydroxylation e.g. morphine, aminopyrine, benzpyrineand aniline.
drug-H + O2+ XH2 drug-OH + H2O + X→ drug-OH + H2O + X
Function of mitochondrial cytochromeP450
1-It has a role in biosynthesis of steroid hormones from cholesterol in adrenal cortex, testis, ovary and placenta by hydroxylation
2-It has a role in biosynthesis of bile acids from cholesterol in the liver by hydroxylation at C26 by 26 hydroxylase.
3-It is important for activation of vitamin D
CytochromeP450
It is a group of hydroxylaseswhich are collectively referred to as cytochromeP450.
They are so called because their reduced forms exhibit an intense absorption band at wavelength 450 nm when complexedto carbon monoxide.
They are conjugated protein containing haeme(haemoproteins).
According to their intracellular localization they may be classified into:
•MicrosomalcytochromeP450.
It is present mainly in the microsomes of liver cells. It represents about 14% of the microsomalfraction of liver cells.
•Mitochondrial cytochromeP450.
It is present in mitochondria of many tissues but it is particularly abundant in liver and steroidogenictissues as adrenal cortex, testis, ovary, placenta and kidney.
Tissue Respiration.
Tissue respiration is the release of energy, usually from glucose, in the tissues of all animals, green plants, fungi and bacteria. All these living things require energy for other processes such as growth, movement, sensitivity, and reproduction.
The most efficient form of respiration is aerobic respiration: this requires oxygen. When oxygen is not available, some organisms can respire anaerobically i.e. without air or oxygen. Yeast can respire in both ways. Yeast gets more energy from aerobic respiration, but when it runs out of oxygen it does not die. It can continue to respire anaerobically, but it does not get so much energy from the sugar. Yeast produces ethanol (alcohol) when it respires anaerobically and ultimately the ethanol will kill the yeast.
We can respire in both ways too. Normally we use oxygen, but when we are running in a race, we may not get enough oxygen into our blood, so our muscles start to respire anaerobically. Unlike yeast we produce lactic acid. Of course if we produced alcohol in our muscles it would make us drunk! Fine thing if you are running away from a predator and you end up drunk! Making lactic acid is not much better. Lactic acid causes cramp.
Glucose + Oxygen = Carbon Dioxide + Water + Energy
This word equation means: “sugar and oxygen are turned into carbon dioxide and water releasing energy”. You must memorise the word equation (and the balanced chemical equation if you want a grade A, B or C). Get help memorising the equations
Glucose = Carbon Dioxide + Ethanol + Energy
This word equation means: “glucose is turned into carbon dioxide and ethanol releasing energy”. You must memorise this word equation.
Composition of respiratory chain
NADH dehydrogenase (EC 1.6.5.3) (also referred to as "NADH:quinone reductase" or "Complex I") is an enzyme located in the inner mitochondrial membrane that catalyzes the transfer of electrons from NADH to coenzyme Q (CoQ). It is the "entry enzyme" of oxidative phosphorylation in the mitochondria.
Function
NADH Dehydrogenase is the first enzyme (Complex I) of the mitochondrial electron transport chain. There are three energy-transducing enzymes in the electron transport chain - NADH dehydrogenase (Complex I), Coenzyme Q – cytochrome c reductase (Complex III), and cytochrome c oxidase (Complex IV). NADH dehydrogenase is the largest and most complicated enzyme of the electron transport chain..
The reaction of NADH dehydrogenase is:
In this process, the complex translocates four protons across the inner membrane per molecule of oxidized NADH, helping to build the electrochemical potential used to produce ATP.
The reaction can be reversed - referred to as aerobic succinate-supported NAD+ reduction - in the presence of a high membrane potential, but the exact catalytic mechanism remains unknown.
Complex I may have a role in triggering apoptosis. [5] In fact, there has been shown to be a correlation between mitochondrial activities and programmed cell death (PCD) during somatic embryo development.
Mechanism
All redox reactions take place in the extramembranous portion of NADH dehydrogenase. NADH initially binds to NADH dehydrogenase, and transfers two electrons to the flavin mononucleotide (FMN) prosthetic group of complex I, creating FMNH2. The electron acceptor - the isoalloxazine ring - of FMN is identical to that of FAD. The electrons are then transferred through the second prosthetic group of NADH dehydrogenase via a series of iron-sulfur (Fe-S) clusters, and finally to coenzyme Q (ubiquinone). This electron flow causes four hydrogen ions to be pumped out of the mitochondrial matrix. Ubiquinone (CoQ) accepts two electrons to be reduced to ubiquionol (CoQH2). [2]
FADH dehydrogenase
In biochemistry, flavin adenine dinucleotide (FAD) is a redox cofactor involved in several important reactions in metabolism. FAD can exist in two different redox states, which it converts between by accepting or donating electrons. The molecule consists of a riboflavin moiety (vitamin B2) bound to the phosphate group of an ADP molecule. The flavin group is bound to ribitol, a sugar alcohol, by a carbon-nitrogen bond, not a glycosidic bond. Thus, riboflavin is not technically a nucleotide; the name flavin adenine dinucleotide is a misnomer.[1]
FAD can be reduced to FADH2, whereby it accepts two hydrogen atoms (a net gain of two electrons):

FAD (fully oxidized form, or quinone form) accepts two electrons and two protons to become FADH2 (hydroquinone form). FADH2 can then be oxidized to the semireduced form (semiquinone) FADH by donating one electron and one proton. The semiquinone is then oxidized once more by losing an electron and a proton and is returned to the initial quinone form (FAD).
FAD is an aromatic ring system, whereas FADH2 is not. This means that FADH2 is significantly higher in energy, without the stabilization that aromatic structure provides. FADH2 is an energy-carrying molecule, because, if it is oxidized, it will regain aromaticity and release all the energy represented by this stabilization.
The primary biochemical role of FADH2 in eukaryotes is to carry high-energy electrons used for oxidative phosphorylation. Its hydrogens remain in the mitochondrial matrix, whilst FAD is tightly bound to a dehydrogenase enzyme i.e. the second protein complex in the oxidative phosphorylation chain. FAD is a prosthetic group in the enzyme complex succinate dehydrogenase (complex II) that oxidizes succinate to fumarate in the eighth step of the citric acid cycle. The high-energy electrons from this oxidation are stored momentarily by reducing FAD to FADH2. FADH2 then reverts to FAD, sending its two high-energy electrons through the electron transport chain; the energy in FADH2 is enough to produce 1.5 equivalents of ATP[2] by oxidative phosphorylation. Another metabolic source of FADH2 is beta oxidation, where FAD serves as a coenzyme to acyl CoA dehydrogenase. A flavoprotein is a protein that contains a flavin moiety, this may be in the form of FAD or FMN (Flavin mononucleotide) . There are many flavoproteins besides components of the succinate dehydrogenase complex, including α-ketoglutarate dehydrogenase and a component of the pyruvate dehydrogenase complex.
Ubiquinones
These molecules are also known as coenzyme Q or mitoquinones. They are involved in electron transport in mitochondrial preparations playing an essential role in the oxidation of succinate or NADH via the cytochrome system. They serves not only as a coenzyme but also, in their reduced forms, as antioxidants. They are synthesized de novo in all animal tissues and cannot thus be regarded as vitamins. Ubiquinones are present in all aerobic organisms, plants, animals (the name ubiquinone was proposed with reference to their ubiquitous occurrence) and bacteria, but are absent from Gram-positive eubacteria and the archaebacteria. They were discovered by the Morton's group in animal fat but their quinonoid structure was revealed by Crane two years later in extracts from beef heart mitochondria.
The compound had a 2,3-dimethoxy-5-methylbenzoquinone nucleus and a side chain of 10 isoprenoid units and was referred to as coenzyme Q 10 . Later, homologues with 6, 7, 8 and 9 units were isolated from other organisms, bacteria or higher organisms. The main form in man has 10 units but in rat has 9 units. Another system of nomenclature is used: ubiquinone(x) in which x designates the total number of carbon atoms in the side chain, it can be a multiple of 5.
Ubiquinones accept one electron and are transformed into semiquinone radicals (UQH°) or two electrons to give ubiquinol (UQH2)
Coenzyme Q is reducible by sodium dithionite or borohydride to its hydroquinone form, and can in turn be reoxidized to the quinone by Ag2O or more slowly by oxygen. The absorption spectra of the two forms are shown below. The quinone form has a strong absorption band at 275 nm which disappears in the reduced form.
Cytochromes are, in general, membrane-bound (i.e. inner mitochondrial membrane) hemeproteins containing heme groups and are primarily responsible for the generation of ATP via electron transport.
They are found either as monomeric proteins (e.g., cytochrome c) or as subunits of bigger enzymatic complexes that catalyze redox reactions.
History
Cytochromes were initially described in 1884 by MacMunn as respiratory pigments (myohematin or histohematin).[1] In the 1920s, Keilin rediscovered these respiratory pigments and named them the cytochromes, or “cellular pigments”, and classified these heme proteins, on the basis of the position of their lowest energy absorption band in the reduced state, as cytochromes a (605 nm), b (~565 nm), and c (550 nm). The UV-visible spectroscopic signatures of hemes are still used to identify heme type from the reduced bis-pyridine-ligated state, i.e., the pyridine hemochrome method. Within each class, cytochrome a, b, or c, early cytochromes are numbered consecutively, e.g. cyt c, cyt c1, and cyt c2, with more recent examples designated by their reduced state R-band maximum, e.g. cyt c559.[2]
Structure and function
The heme group is a highly-conjugated ring system (which allows its electrons to be very mobile) surrounding a metal ion, which readily interconverts between the oxidation states. For many cytochromes, the metal ion present is that of iron, which interconverts between Fe2+ (reduced) and Fe3+ (oxidised) states (electron-transfer processes) or between Fe2+ (reduced) and Fe3+ (formal, oxidized) states (oxidative processes). Cytochromes are, thus, capable of performing oxidation and reduction. Because the cytochromes (as well as other complexes) are held within membranes in an organized way, the redox reactions are carried out in the proper sequence for maximum efficiency.
In the process of oxidative phosphorylation, which is the principal energy-generating process undertaken by organisms, other membrane-bound and -soluble complexes and cofactors are involved in the chain of redox reactions, with the additional net effect that protons (H+) are transported across the mitochondrial inner membrane. The resulting transmembrane proton gradient ([protonmotive force]) is used to generate ATP, which is the universal chemical energy currency of life. ATP is consumed to drive cellular processes that require energy (such as synthesis of macromolecules, active transport of molecules across the membrane, and assembly of flagella).
Types
Several kinds of cytochrome exist and can be distinguished by spectroscopy, exact structure of the heme group, inhibitor sensitivity, and reduction potential.
Three types of cytochrome are distinguished by their prosthetic groups:
|
Type |
Prosthetic group |
The definition of cytochrome c is not defined in terms of the heme group. There is no "cytochrome e," but there is a cytochrome f, which is often considered a type of cytochrome c.
In mitochondria and chloroplasts, these cytochromes are often combined in electron transport and related metabolic pathways:
|
Cytochromes |
Combination |
|
a and a3 |
Cytochrome c oxidase ("Complex IV") with electrons delivered to complex by soluble cytochrome c (hence the name) |
|
b and c1 |
Coenzyme Q - cytochrome c reductase ("Complex III") |
|
b6 and f |
A completely distinct family of cytochromes is known as the cytochrome P450 oxidases, so named for the characteristic Soret peak formed by absorbance of light at wavelengths near 450 nm when the heme iron is reduced (with sodium dithionite) and complexed to carbon monoxide. These enzymes are primarily involved in steroidogenesis and detoxification.
Respiratory chain
The system of mitochondrial enzymes and redox carrier molecules which ferry reducing equivalents from substrates to oxygen are collectively known as the electron transport system, or the respiratory chain. This system captures the free energy available from substrate oxidation so that it may later be applied to the synthesis of ATP. Many respiratory chain components were first identified in crude homogenates through their spectral properties, which frequently change when a carrier is oxidised or reduced. Fractionation of mitochondria in the presence of mild detergents or chaotropic salts dissected the respiratory chain into four large multi-subunit complexes containing the principal respiratory carriers, named Complex 1 to Complex 4.
These substantial protein "icebergs" float in the sheet of inner membrane lipids, often presenting one face to the mitochondrial matrix and another to the inter - membrane space. Many of their components have now been isolated in a relatively pure form. Other membrane bound enzymes such as the energy linked transhydrogenase (ELTH) are also present which fulfil ancillary roles.
The main components participate in the approximate order of their redox potentials, and the bulky complexes are linked by low molecular weight mobile carriers which ferry the reducing equivalents from one complex to the next. Except for succinate dehydrogenase (complex 2) all these complexes pump protons from the matrix space into the cytosol as they transfer reducing equivalents (either hydrogen atoms or electrons) from one carrier to the next. The diagram above shows the flow of reducing equivalents in purple, and movement of the positively charged protons in red. Proton pumping is an arduous task which creates substantial pH and electrical gradients across the mitochondrial inner membrane. These protons eventually re-enter the matrix space via the F1 ATPase, driving the synthesis of ATP as they return.
The number of protons and the number of positive charges crossing the inner membrane need not necessarily agree for each individual transmembrane protein, although the accounts must balance for the whole ensemble. This discrepancy is illustrated on the diagram above, and is explained in greater detail below.
Electrons flow through the electron transport chain to molecular oxygen; during this flow, protons are moved across the inner membrane from the matrix to the intermembrane space. This model for ATP synthesis is called the chemiosmotic mechanism, or Mitchell hypothesis. Peter Mitchell, a British biochemist, essentially by himself and in the face of contrary opinion, proposed that the mechanism for ATP synthesis involved the coupling between chemical energy (ATP) and osmotic potential (a higher concentration of protons in the intermembrane space than in the matrix). The inner membrane of the mitochondrion is tightly packed with cytochromes and proteins capable of undergoing redox changes. There are four major protein-membrane complexes.
Complex I and Complex II
Complex I and Complex II direct electrons to coenzyme Q. Complex I, also called NADH-coenzyme Q reductase, accepts electrons from NADH. The NADH releases a proton and two electrons. The electrons flow through a flavoprotein containing FMN and an iron-sulfur protein. First, the flavin coenzyme (flavin mononucleotide) and then the iron-sulfur center undergo cycles of reduction and then oxidation, transferring their electrons to a quinone molecule, coenzyme Q (see Figure ).
Complex I is capable of transferring protons from the matrix to the intermembrane space while undergoing these redox cycles. One possible source of the protons is the release of a proton from NADH as it is oxidized to NAD, although this is not the only explanation. Apparently, conformational changes in the proteins of Complex I also are involved in the mechanism of proton translocation during electron transport.
|
|
Complex II, also known as succinate-coenzyme Q reductase, accepts electrons from succinate formed during the TCA cycle. Electrons flow from succinate to FAD (the flavin-adenine dinucleotide) coenzyme, through an iron-sulfur protein and a cytochrome b550 protein (the number refers to the wavelength where the protein absorbs), and to coenzyme Q. No protons are translocated by Complex II. Because translocated protons are the source of the energy for ATP synthesis, this means that the oxidation of a molecule of FADH2 inherently leads to less ATP synthesized than does the oxidation of a molecule of NADH. This experimental observation also fits with the difference in the standard reduction potentials of the two molecules. The reduction potential of FAD is -0.22 V, as opposed to -0.32 V for NAD.
Coenzyme Q is capable of accepting either one or two electrons to form either a semiquinone or hydroquinone form. Coenzyme Q is not bound to a protein; instead it is a mobile electron carrier and can float within the inner membrane, where it can transfer electrons from Complex I and Complex II to Complex III.
Complex III is also known as coenzyme Q-cytochrome c reductase. It accepts electrons from reduced coenzyme Q, moves them within the complex through two cytochromes b, an iron-sulfur protein, and cytochrome c1. Electron flow through Complex II transfers proton(s) through the membrane into the intermembrane space. Again, this supplies energy for ATP synthesis. Complex III transfers its electrons to the heme group of a small, mobile electron transport protein, cytochrome c.
Cytochrome c transfers its electrons to the final electron transport component, Complex IV, or cytochrome oxidase. Cytochrome oxidase transfers electrons through a copper-containing protein, cytochrome a, and cytochrome a3, and finally to molecular oxygen. The overall pathway for electron transport is therefore:
|
||
|
|
|
The number n is a fudge factor to account for the fact that the exact stoichiometry of proton transfer isn't really known. The important point is that more proton transfer occurs from NADH oxidation than from FADH2 oxidation.
Chemiosmotic hypothesis
A theory postulated by the biochemist Peter Mitchell in 1961 to describe ATP synthesis by way of a proton electrochemical coupling is called chemiosmotic hypothesis.
Accordingly, hydrogen ions (protons) are pumped from the mitochondrial matrix to the intermembrane space via the hydrogen carrier proteins while the electrons are transferred along the electron transport chain in the mitochondrial inner membrane. As the hydrogen ions accumulate in the intermembrane space, an energy-rich proton gradient is established. As the proton gradient becomes sufficiently intense the hydrogen ions tend to diffuse back to the matrix (where hydrogen ions are less) via the ATP synthase (a transport protein). As the hydrogen ions diffuse (through the ATP synthase) energy is released which is then used to drive the conversion of ADP to ATP (by phosphorylation).
Chemiosmotic Hypothesis in a simple form
In the 1960s, ATP was known to be the energy currency of life, but the mechanism by which ATP was created in the mitochondria was assumed to be by substrate-level phosphorylation. Mitchell's chemiosmotic hypothesis was the basis for understanding the actual process of oxidative phosphorylation. At the time, the biochemical mechanism of ATP synthesis by oxidative phosphorylation was unknown.
Mitchell realised that the movement of ions across an electrochemical membrane potential could provide the energy needed to produce ATP. His hypothesis was derived from information that was well known in the 1960s. He knew that living cells had a membrane potential; interior negative to the environment. The movement of charged ions across a membrane is thus affected by the electrical forces (the attraction of positive to negative charges). Their movement is also affected by thermodynamic forces, the tendency of substances to diffuse from regions of higher concentration. He went on to show that ATP synthesis was coupled to this electrochemical gradient.
His hypothesis was confirmed by the discovery of ATP synthase, a membrane-bound protein that uses the potential energy of the electrochemical gradient to make ATP.
The passage back occurs via a specific proton channel. This passage is coupled to ATP-synthesis, using the potential energy of the proton gradient for the formation of the third phosphate bond of ATP.
We can now calculate the end result of glucose degradation: the oxidation is coupled to a decrease of the free energy; 686 kcal/mol (= 2881 kJ/mol) are obtained by the complete oxidation of glucose. How much of this energy can the cell use?
1. Six mol ATP per mol glucose are generated (substrate chain phosphorylation). This is because all steps after the breaking down of fructose-1,6-phosphate have to be counted twice (once for each of the two resulting C3 molecules), so it is 3 x 2 ATPs. Of these six ATPs, two are needed to start glycolysis. That leaves four.
2. During the course of glycolysis up to acetyl-CoA, 2 x 2 NADH + H+ are generated. An additional 3 x 2 NADH + H+ and 1 x 2 FADH2 are produced in the citric acid cycle. One NADH + H+ gives three, one FADH2 two ATPs when fed into the respiratory chain. This sums up to 34 ATPs plus the 4 ATPs of glycolysis. A total of 38 mol ATP are thus gained by the cell's degradation of one mol glucose. Since each energy-rich bond of ATP contains 7.3 kcal/mol (= -30.6 kJ/mol), the 38 ATP equal 277 kcal/mol (ca 1163 kJ/mol). This is 40.6% of the theoretically possible gain. The other 59.4 percent are set free as heat. This is a very high percentage compared to the gain of technical machines like steam or petrol engines that is around or below 20 percent.
The ATP synthase enzymes have been remarkably conserved through evolution. The bacterial enzymes are essentially the same in structure and function as those from mitochondria of animals, plants and fungi, and the chloroplasts of plants. The early ancestory of the enzyme is seen in the fact that the Archaea have an enzyme which is clearly closely related, but has significant differences from the Eubacterial branch. The H+-ATP-ase found in vacuoles of the eukaryote cell cytoplasm is similar to the archaeal enzyme, and is thought to reflect the origin from an archaeal ancestor.
In most systems, the ATP synthase sits in the membrane (the "coupling" membrane), and catalyses the synthesis of ATP from ADP and phosphate driven by a flux of protons across the membrane down the proton gradient generated by electron transfer. The flux goes from the protochemically positive (P) side (high proton electrochemical potential) to the protochemically negative (N) side. The reaction catalyzed by ATP synthase is fully reversible, so ATP hydrolysis generates a proton gradient by a reversal of this flux. In some bacteria, the main function is to operate in the ATP hydrolysis direction, using ATP generated by fermentative metabolism to provide a proton gradient to drive substrate accumulation, and maintain ionic balance.
ADP + Pi + nH+P <=> ATP + nH+N
- the free energy change (DG) associated with synthesis of ATP under cellular conditions (the free energy required)
Because the structures seen in EM, the subunit composition, and the sequences of the subunits appeared to be so similar, it had been assumed that the mechanisms, and hence the stoichiometries, would be the same. In this context, the evidence suggesting that the stoichiometry of H+/ATP (n above) varied depending on system was surprising. Values based on measure ATP/2e- ratios, and H+/2e- ratios had suggested that n was 3 for mitochondria, and 4 for chloroplasts, but these values were based on the assumption of integer stoichiometries. Although all the F1F0-type ATP-synthases likely had a common origin, both the assumption that the stoichiometries are the same, and that n is integer, are called into question by emerging structural data (see below).
Structure of the F1 ATP-ase
The structure of the soluble (F1) portion of the ATP synthase from beef heart mitochondria has been solved by X-ray crystallography. The pictures below are from Abrahams, J.P., Leslie, A.G., Lutter, R. and Walker, J.E. (1994) Structure at 2.8 Å resolution of F1-ATPase from bovine heart mitochondria
Mechanism of the F1 ATP-ase
The ATP synthase operates through a mechanism in which the three active sites undergo a change in binding affinity for the reactants of the ATP-ase reaction, ATP, ADP and phosphate, as originally predicted by Paul Boyer. The change in affinity accompanies a change in the position of the g-subunit relative to the a, b-ring, which involves a rotation of the one relative to the other. In the direction of ATP synthesis, the rotation is driven by a flux of H+ down the proton gradient, through a coupling between the g-subunit, and the c-subunit of FO. This rotation has now been demonstrated experimentally.
Respiratory control
The dependence of oxidative phosphorylation on ADP reveals an important general feature of this process: Respiration is tightly coupled to the synthesis of ATP. Not only is ATP synthesis absolutely dependent on continued electron flow from substrates to oxygen, but electron flow in normal mitochondria occurs only when ATP is being synthesized as well. This regulatory phenomenon, called respiratory control, makes biological sense, because it ensures that substrates will not be oxidized wastefully. Instead, their utilization is controlled by the physiological need for ATP.
In most aerobic cells the level of ATP exceeds that of ADP by 4- to 10-fold. Respiration depends on ADP as a substrate for phosphorylation. When ATP is consumed at high rates, accumulation of ADP stimulates respiration, with concomitant activation of ATP resynthesis. Conversely, in a relaxed and well-nourished cell, ATP accumulates at the expense of ADP, and the depletion of ADP limits the rate of both electron transport and its own phosphorylation to ATP. Thus, the energy-generating capacity of the cell is closely attuned to its energy demands.
Experimentally, respiratory control is demonstrated by following oxygen utilization in isolated mitochondria. In the absence of added substrate or ADP, oxygen uptake, caused by oxidation of endogenous substrates, is slow. Addition of an oxidizable substrate, such as glutamate or malate, has but a small effect on the respiration rate. If ADP is then added, however, oxygen uptake proceeds at an enhanced rate until all of the added ADP has been converted to ATP, and then oxygen uptake returns to the basal rate. This stimulation of respiration is stoichiometric; that is, addition of twice as much ADP causes twice the amount of oxygen uptake at the enhanced rate. If excess ADP is present instead of oxidizable substrate, the addition of substrate in limiting amounts will stimulate oxygen uptake until the substrate is exhausted.
Two mechanisms of the control of respiration and ATP synthesis in mitochondria according to the utilization of energy (ATP). The first mechanism of respiratory control is based on the proton motive force Δp across the inner mitochondrial membrane (grey). Activation of the ATP-synthase (blue) by ADP, taken up via the ATP/ADP carrier (margenta), decreases Δp which in consequence stimulates the three proton pumps of the respiratory chain (complexes I, III and IV). For simplicity, only complex IV (cytochrome c oxidase) and its substrate (cytochrome c) are shown in green and red, respectively. The second mechanism of respiratory control is based on the intramitochondrial ATP/ADP ratio. High ATP/ADP ratios inhibit cytochrome c oxidase activity allosterically. Uptake of ADP decreases the intramitochondrial ATP/ADP ratio accompanied by exchange of bound ATP by ADP at the matrix domain of subunit IV of cytochrome c oxidase, with subsequent stimulation of respiration
Maintenance of respiratory control depends on the structural integrity of the mitochondrion. Disruption of the organelle causes electron transport to become uncoupled from ATP synthesis. Under these conditions, oxygen uptake proceeds at high rates even in the absence of added ADP. ATP synthesis is inhibited, even though electrons are being passed along the respiratory chain and used to reduce O2 to water.
Uncoupling of respiration from phosphorylation can also be achieved chemically. Chemical uncouplers such as DNP or FCCP act by dissipating the proton gradient. Addition of an uncoupler to mitochondria stimulates oxygen utilization even in the absence of added ADP. No phosphorylation occurs under these conditions because there is no ADP to be phosphorylated.
The phenomenon of respiratory control is the subject of today's studio exercise. An oxygen electrode may be used to record [O2] in a closed vessel (diagram p. 804). Electron transfer, e.g., from NADH to O2, is monitored by recording the rate of disappearance of O2. At right is an idealized representation of an oxygen electrode recording while mitochondria respire in the presence of Pi, along with an electron donor (e.g., succinate, or a substrate of a reaction that will generate NADH). The dependence of respiration rate on availability of ADP, the substrate for the ATP Synthase, is called respiratory control. The respiratory control ratio is the ratio of slopes after and before ADP addition (b/a).
The P/O ratio is the moles of ADP added, divided by the moles of O consumed (based on c) while phosphorylating the added ADP.
Chemiosmotic explanation of respiratory control:
Electron transfer is obligatorily coupled to H+ ejection from the matrix. Whether this coupled reaction is spontaneous depends on the pH and electrical gradients.
|
Reaction |
Free energy change |
|
e- transfer (e.g., NADH to O2) |
a negative value* |
|
H+ ejection from the matrix |
a positive value that varies with the H+ gradient** |
|
e- transfer coupled to H+ ejection |
algebraic sum of the above |
*DGo' = - nFDEo' = -218 kJ/mol, for transfer of 2 e- from NADH to O2.
** For ejection of one H+
from the matrix:
DG = RT ln ([H+]cytosol/[H+]matrix)
+ F DY = 2.3 RT (pHmatrix - pHcytosol) + F DY
In the absence of ADP, H+ cannot flow back to the matrix through Fo. The pH and electrical gradients (DpH & DY) are maximal. As respiration with outward H+ pumping proceeds, the free energy change for H+ ejection (positive DG) increases and approaches the magnitude of that for electron transfer (negative DG). When the coupled reaction becomes non-spontaneous, respiration stops. This is referred to as a static head. In fact there is usually a low rate of respiration in the absence of ADP, attributed to H+ leaks. Protons pumped out are carried by the uncoupler back into the mitochondrial matrix, preventing development of a pH or electrical gradient.
Inhibitors of tissue respiration
Chemiosmosis can be disrupted by a variety of chemicals. In oxidative phosphorylation, some of these inhibitors are quite infamous:
The electron transport chain was determined by studying the effects of particular inhibitors.
Rotenone is a common insecticide that strongly inhibits the electron transport of complex I.
Rotenone is a natural product obtained from the roots of several species of plants. Tribes in certain parts of the world beat the roots of trees along riverbanks to release rotenone into the water which paralyzes fish and makes them easy prey.
Amytal
is a barbiturate that inhibits the electron transport of complex I. Demerol is
painkiller that also inhibits complex
Antibiotic. Induces apoptosis, which is not prevented by the presence of Bcl-2. Inhibits mitochondrial electron transport specifically between cytochromes b and c1. All three of these complex I inhibitors block the oxidation of the Fe-S clusters of complex I.
2-Thenoyltrifluoroacetone and carboxin specifically block electron transport in Complex II
Antimycin A is an antibiotic that inhibits electron transfer in complex III by blocking the transfer of electrons between Cyt bH and coenzyme Q bound at the QN site. Antibiotic. Induces apoptosis, which is not prevented by the presence of Bcl-2. Inhibits mitochondrial electron transport specifically between cytochromes b and c1.
Cyanide, azide and carbon monoxide all inhibit electron transport in Complex IV. The all inhibit electron transfer by binding tightly with the iron coordinated in Cyt a
This complex oxidizes cytochrome c and also reduces O2 to H2O. Remember that cytochromes have heme cofactors -- this is important in our discussion of cyanide and azide. Cytochrome c is a soluble protein and also is a mobile carrier. Other inhibitors of cytochrome c oxidase will not be discussed here, but are important biologically, such as sulfide, formate, and nitric oxide.
Azide and cyanide bind to the iron when the iron is in the ferric state. Carbon Monoxide binds to the iron when it is in the ferrous state. Cyanide and azide are potent inhibitors at this site which accounts for there acute toxicity. Carbon monoxide is toxic due to its affinity for the heme iron of hemoglobin. Animals carry many molecules of hemoglobin, therefore it takes a large quantity of carbon monoxide to die from carbon monoxide poisoning.
Carbon Monoxide
CO competes with oxygen for binding to the reduced form of cytochrome c oxidase. Once bound to the cytochrome oxidase, oxygen cannot attach, and electron transport is stopped. CO is a colorless, tasteless, non-irritating toxic gas. When inhaled, the toxic gas enters the bloodstream, depriving the heart and brain of the oxygen necessary to function correctly. Sensing the body's need for more oxygen, the victim's heart rate increases to pump more blood to the body's organs. If a person continues to inhale CO, he or she faces the risk of breathing difficulty, cardiac trauma, brain damage, coma and even death.
Animals have relatively few molecules of Cyt a3. Consequently an exposure to a small quantity of azide or cyanide can be lethal. The toxicity of cyanide is solely from its ability to arrest electron transport. Cyanide
CN-, like azide, binds to the iron atom of oxidized cytochrome, preventing binding of oxygen. Again, since cytochrome oxidase is inhibited, oxygen metabolism is prevented and thus so is energy generation. Cyanide has long been known as a poison, sometimes used in warfare. As mentioned above, lethal doses cause death in 15 minutes.
How does cyanide, azide or carbon monoxide cause death?
They
all cause similar toxic repercussions (with the exception of Carbon Monoxide,
which can bind to Hemoglobin causing your body to be unable to bind oxygen
properly and causing you to suffocate);
However, these three also cause chemical suffocation. They bind a protein in
our electron transport chain (cytochrome c oxidase). The electron transport
chain is our body's best way to create energy in the form of ATP. Without it,
we literally have next to no energy. The electron transport chain is the reason
we have pain in our muscles when we work out--if our body doesn't have oxygen
in those tissues and can't make energy, we make lactic acid, which causes our
muscles to hurt/burn. Anyway, cyanide, azide and carbon monoxide bind this
protein (cytochrome c oxidase) in the electron transport chain. Doing this
causes all of the electrons to stop transferring and no energy to be made. This
causes cell death. The first place to experience cell death from cyanide, azide
and carbon monoxide is your central nervous system--which is made up of your
brain and spinal cord.
So the first thing to die when someone dies of cyanide, azide or carbon
monoxide poisoning?
The coupling between electron transport and oxidative phosphorylation depends on the impermeability of the inner mitochondrial membrane to H+translocation. The only way for protons to go from the intermembrane space to the matrix is through ATP synthase. Uncouplers uncouple electron transport from oxidative phosphorylation. They collapse the chemiosmotic gradient by dissipating protons across the inner mitochondrial membrane. All of the uncouplers shown to the left, collapse the pH gradient by binding a proton on the acidic side of the membrane, diffusing through the inner mitochondrial membrane and releasing the proton on the membranes alkaline side.
Uncouplers of oxidative phosphorylation stimulate the rate of electron flow but not
ATP synthesis.
(a)At relatively low levels of an uncoupling agent, P/O ratios drop somewhat, but the cell can compensate for this by increasing the rate of electron flow; ATP levels can be kept relatively normal. At high levels of uncoupler, P/O ratios approach zero and the cell cannot maintain ATP levels.
(b) As amounts of an uncoupler increase, the P/O ratio decreases and the body struggles to make sufficient ATP by oxidizing more fuel. The heat produced by this increased rate of oxidation raises the body temperature. The P/O ratio is affected as noted in (a).
(c) Increased activity of the respiratory chain in the presence of an uncoupler requires the degradation of additional energy stores (glycogen and fat). By oxidizing more fuel in an attempt to produce the same amount of ATP, the organism loses weight. If the P/O ratio nears zero, the lack of ATP will be lethal
2,4-Dinitrophenol, dicumarol and carbonyl cyanide-p-trifluorocarbonyl-cyanide methoxyphenyl hydrazone (FCCP) all have hydrophobic character making them soluble in the bilipid membrane. All of these decouplers also have dissociable protons allowing them to carry protons from the intermembrane space to the matrix which collapses the pH gradient. The potential energy of the proton gradient is lost as heat DNP is a chemical uncoupler of electron transport and oxidative phosphorylation.
DNP permeabilizes the inner mitochondrial membrane to protons, destroying the proton gradient and, in doing so, uncouples the electron transport system from the oxidative phosphorylation. In this situation, electrons continue to pass through the electron transport system and reduce oxygen to water, but ATP is not synthesized in the process. The compound, trifluorocarbonylcyanide phenylhydrazone (FCCP), is also an uncoupler.
The phenolic group of DNP is usually dissociated at intracellular pH. However, a DNP molecule that approaches the inner mitochondrial membrane from the outside becomes protonated (because the pH is lower there). Protonation increases the hydrophobicity of DNP, allowing it to diffuse into the membrane and, by mass action, to pass through. Once inside, the higher pH of the matric deprotonates the phenolic hydroxyl again. Thus, DNP has the effect of transporting H+ back into the matrix, bypassing the F0 proton channel and thereby preventing ATP synthesis.
The link between electron transport and ATP synthesis is below. (a) In the presence of excess phosphate and substrate and intact mitochondria, oxygen is consumed only when ADP is added. When all of the added ADP has been converted into ATP, electron transport stops and oxygen consumption ceases. (b) The addition of 2,4-dintrophenol uncouples electron transfer from ATP synthesis. The oxygen is completely consumed in the absence of ADP. Endogenous Uncouplers Enable Organisms to Generate Heat
.
The uncoupling of oxidative phosphorylation from electron transport generates heat. Hibernating animals and newborne animals (including human beings) contain brown adipose tissue. The adipose tissue is brown due to the high mitochondria content of the tissue. An endogenous protein called thermogenin uncouples ATP synthesis from electron transport by opening up a passive proton channel (UCP-1) through the inner mitochondrial membrane. The collapse of the pH gradient generates heat. An uncoupling protein (also called thermogenin) is produced in brown adipose tissue of newborn mammals and hibernating mammals .
This protein of the inner mitochondrial membrane functions as a H+ carrier. The uncoupling protein blocks development of a H+ electrochemical gradient, thereby stimulating respiration. The free energy change associated with respiration is dissipated as heat. This "non-shivering thermogenesis" is costly in terms of respiratory energy unavailable for ATP synthesis, but it provides valuable warming of the organism.
Valinomycin
combines with K ions to form a complex that passes through the inner
Valinomycin is a potent antibiotic which acts as a potassium (K+) ionophore. Induces K+ conductivity in cell membranes. Also active in vitro against Mycobacterium Tuberculosis, and as an apoptosis inducer.
Valinomycin is obtained from the cells of several Streptomyces strains, among which "S. tsusimaensis" and S. fulvissimus. It is a member of the group of natural neutral ionophores because it does not have a residual charge. It consists of enantiomers D- and L-valine (Val), D-hydroxyvaleric acid and L-lactic acid. Structures are alternately bound via amide and ester bridges. Valinomycin is highly selective for potassium ions over sodium ions within the cell membrane. It functions as a potassium-specific transporter and facilitates the movement of potassium ions through lipid membranes "down" an electrochemical potential gradient. The stability constant K for the potassium-valinomycin complex is 106 and for the sodium-valinomycin complex only 10. This difference is important for maintaining the selectivity of valinomycin for the transport of potassium ions (and not sodium ions) in biological systems.
Oligomycin is a natural antibiotic isolated from Streptomyces
diastatochromogenes which inhibits mitochondrial H+-ATP
synthase. It is primarily found to act as an inhibitor of mitochondrial
respiration and swelling. This antibiotic is widely used as an
inhibitor of oxidative phosphorylation.1-2
Oligomycin inhibits the H+- ATP-synthase by binding to the
Oligomycin sensitivity-conferring protein (OSCP) at the F(o)
subunits 6 and 9 which are found in the stalk of the F1F0-ATPase
complex. This binding blocks the proton conductance and inhibits the synthesis
of mitochondrial ATP.3-4 Because of its activity,
it can also be used to reduce the number of parameters (such as ER Ca2+
release, exocytotoxicity and apopotosis) which are affected by mitochondrial
depolarization.
Oligomycin, at high concentrations may also inhibit the plasma membrane
Na+-K+-ATPase. Interaction of Oligomycin with the Na+
occlusion site on the extracellular side of Na/K-ATPase, delays Na+
release to the extracellular side without inducing a conformational change.
Although Oligomycin stimulated Na+ binding to Na+/K+-ATPase,
it inhibited Na+/Na+ exchange, and did not affect
either Na+-dependent AD/-ATP exchange or K+-dependent
phosphatase activity.6-16
Proton Motive Force Drives Transport The primary purpose of the proton gradient is togenerate ATP by oxidative phosphorylation. The potential energy of the gradient can also be used for active transport. The inner mitochondrial membrane is impermeable to charged molecules.
ATP-ADP Translocase There are two specific systems to transport ADP and Pi into the mitochondrial matrix. A specific transport protein ATP-ADP translocase enables ATP and ADP to transverse the inner mitochondrial membrane. The transport of ADP in and ATP out are coupled. ADP only enters the matrix if ATP exits or vice versa. ATP-ADP translocase has a single nucleotide binding site which binds ADP and ATP with equal affinity. Due to the negative electrostatic charge of the matrix, ATP-is bound on the N phase of the membrane because it has greater negative charge than ADP.
Pentachlorophenol (PCP) acts in a similar way to DNP. It was widely used as a biocide, especially in pallet board manufacture as a fungicide, but is now banned by the Biocidal Products Directive, because of its extreme toxicity and environmental persistence.
DNP
PCP
Ways of energy usage in the organism.
o 1. From the smallest, single-celled organism to the biggest and most complex mammals--including people--all living things require energy for life. It's easy enough to understand that we and other animals eat. Things get a little more puzzling when we think about fungi, which absorb their food as organic molecules, from the surrounding environment. Where do those molecules come from? Furthermore, where does the food come from that we humans convert to energy? At the most basic level, all energy traces back to plants. Plants are the basis of all the world's food systems, and their unique ability to make organic materials from sunlight--called photosynthesis--is what sustains nearly every other life form on the planet.
o The powerhouse of energy production in all plants is called a chloroplast. More than a million of these handy devices occur in every quarter-inch of a leaf. They contain the pigment called chlorophyll that makes most leaves green--and drives photosynthesis. The reaction isn't all that complicated, as far as chemical reactions go. The chloroplasts take in carbon dioxide, sunlight and water. They release oxygen and a bit less water than they took in. The conversion of carbon dioxide to oxygen is one life-sustaining function that plants perform for Earth and all of its life. But plants do something equally as important when they keep a third product behind: glucose, the sugar that sustains the plants---and anything, in turn, that eats the plants.
o In cellular respiration, glucose is broken down by the removal of its hydrogen atoms. That process releases energy in the form of electrons, negatively charged particles that fuel all of a cell's other work in later reactions. So, plants make the glucose and everything down the line---from plant-eaters to the carnivores that eat them---break the glucose down again, and use its energy. That's the simple story. Of course, life is rarely so simple, and there are exceptions to every rule. Every so often, a new discovery comes along about living things that use a non-living substance other than sunlight to make energy--like ammonia, or even sulfur. These less-common organisms can harness electrons from chemical sources instead of the sun. More amazing life forms have the potential to be discovered at any time, anywhere on our planet---or beyond.
Cytochrome P450
The active site of cytochrome P450 contains a heme iron center. The iron is tethered to the P450 protein via a thiolate ligand derived from a cysteine residue. This cysteine and several flanking residues are highly conserved in known CYPs and have the formal PROSITE signature consensus pattern [FW] - [SGNH] - x - [GD] - {F} - [RKHPT] - {P} - C - [LIVMFAP] - [GAD].[7] Because of the vast variety of reactions catalyzed by CYPs, the activities and properties of the many CYPs differ in many aspects. In general, the P450 catalytic cycle proceeds as follows:
1. The substrate binds to the active site of the enzyme, in close proximity to the heme group, on the side opposite to the peptide chain. The bound substrate induces a change in the conformation of the active site, often displacing a water molecule from the distal axial coordination position of the heme iron,[8] and sometimes changing the state of the heme iron from low-spin to high-spin.[9] This gives rise to a change in the spectral properties of the enzyme, with an increase in absorbance at 390 nm and a decrease at 420 nm. This can be measured by difference spectrometry and is referred to as the "type I" difference spectrum (see inset graph in figure). Some substrates cause an opposite change in spectral properties, a "reverse type I" spectrum, by processes that are as yet unclear. Inhibitors and certain substrates that bind directly to the heme iron give rise to the type II difference spectrum, with a maximum at 430 nm and a minimum at 390 nm (see inset graph in figure). If no reducing equivalents are available, this complex may remain stable, allowing the degree of binding to be determined from absorbance measurements in vitro
2. The change in the electronic state of the active site favors the transfer of an electron from NAD(P)H via cytochrome P450 reductase or another associated reductase[11] This takes place by way of the electron transfer chain, as described above, reducing the ferric heme iron to the ferrous state.
3. Molecular oxygen binds covalently to the distal axial coordination position of the heme iron. The cysteine ligand is a better electron donor than histidine, which is normally found in heme-containing proteins. As a consequence, the oxygen is activated to a greater extent than in other heme proteins. However, this sometimes allows the iron-oxygen bond to dissociate, causing the so-called "uncoupling reaction", which releases a reactive superoxide radical and interrupts the catalytic cycle.
4. A second electron is transferred via the electron-transport system, from either cytochrome
P450 reductase, ferredoxins, or cytochrome b5, reducing the dioxygen adduct to a negatively charged peroxo group. This is a short-lived intermediate state.
5. The peroxo group formed in step 4 is rapidly protonated twice by local transfer from water or from surrounding amino-acid side-chains, releasing one water molecule, and forming a highly reactive species commonly referred to as P450. This highly reactive intermediate was not "seen in action" until 2010,[12] although it had been studied theoretically for many years. P450 Compound 1 is most likely a iron(IV)oxo (or ferryl) species with an additional oxidizing equivalent delocalized over the porphyrin and thiolate ligands. Evidence for the alternative perferryl iron(V)-oxo [8] is lacking.
6. Depending on the substrate and enzyme involved, P450 enzymes can catalyze any of a wide variety of reactions. A hypothetical hydroxylation is shown in this illustration. After the product has been released from the active site, the enzyme returns to its original state, with a water molecule returning to occupy the distal coordination position of the iron nucleus.
S: An
alternative route for mono-oxygenation is via the "peroxide shunt":
Interaction with single-oxygen donors such as peroxides and hypochlorites can
lead directly to the formation of the iron-oxo intermediate, allowing the
catalytic cycle to be completed without going through steps 2, 3, 4, and
C: If carbon monoxide (CO) binds to reduced P450, the catalytic cycle is interrupted. This reaction yields the classic CO difference spectrum with a maximum at 450 nm.