БИОХИМИЧЕСКИЕ ФУНКЦИИ ПЕЧЕНИ. МЕТАБОЛИЗМ
ПОРФИРИНА: ОБМЕН ЖЕЛЧНЫХ ПИГМЕНТОВ, БИОХИМИЯ ЖЕЛТУХ. БИОТРАНСФОРМАЦИЯ
КСЕНОБИОТИКОВ И ЭНДОГЕННЫХ ТОКСИНОВ В
ПЕЧЕНИ: МИКРОСОМАЛЬНОЕ ОКИСЛЕНИЕ, ЦИТОХРОМ Р-450
ПЕЧЕНЬ
Важнейшее
значение печени в обмене
веществ в первую очередь определяется тем, что она является как бы
большой промежуточной станцией между портальным и общим кругом кровообращения.
РОЛЬ ПЕЧЕНИ В УГЛЕВОДНОМ
ОБМЕНЕ
Основная роль
печени в углеводном обмене заключается в обеспечении постоянства концентрации глюкозы в крови. Это
достигается регуляцией между синтезом и распадом гликогена, депонируемого в
печени.
В печени
синтез гликогена
и его регуляция в основном аналогичны тем процессам, которые протекают в других
органах и тканях,
в частности в мышечной ткани.
Синтез гликогена
из глюкозы
обеспечивает в норме временный резерв углеводов, необходимый для
поддержания концентрации
глюкозы в крови в тех
случаях, если ее содержание значительно уменьшается (например, у человека это
происходит при недостаточном поступлении углеводов с пищей или в
период ночного «голодания»).
Необходимо
подчеркнуть важную роль фермента
глюкокиназы в процессе утилизации глюкозы печенью.
Глюкокиназа, подобно гексокиназе,
катализирует фосфорилирование
глюкозы с образованием
глюкозо-6-фосфата, при этом активность
глюкокиназы в печени почти в 10 раз превышает активность гексокиназы.
Важное
различие между этими двумя ферментами заключается в
том, что глюкокиназа в противоположность гексокиназе имеет высокое
значение КМ для глюкозы
и не ингибируется глюкозо-6-фосфатом.
После приема
пищи содержание глюкозы
в воротной вене резко возрастает: в тех же пределах увеличивается и ее
внутрипеченочная концентрация
. Повышение концентрации
глюкозы в печени вызывает
существенное увеличение активности
глюкокиназы и автоматически увеличивает поглощение глюкозы печенью
(образовавшийся глюкозо-6-фосфат либо затрачивается на синтез гликогена, либо
расщепляется).
Считают, что
основная роль печени – расщепление глюкозы – сводится прежде
всего к запасанию метаболитов-предшественников, необходимых для биосинтеза жирных кислот и
глицерина, и в меньшей
степени к окислению
ее до СО2 и Н2О. Синтезированные в печени триглицериды в
норме выделяются в кровь в составе липопротеинов и
транспортируются в жировую ткань
для более «постоянного» хранения.
В реакциях
пентозофосфатного пути в печени образуется НАДФН, используемый для
восстановительных реакций в процессах синтеза жирных кислот, холестерина и других стероидов. Кроме того, при
этом образуются пентозофосфаты, необходимые для синтеза нуклеиновых кислот.
Наряду с
утилизацией глюкозы
в печени происходит и ее образование. Непосредственным источником глюкозы в печени служит гликоген. Распад гликогена в печени
происходит в основном фосфоролитическим путем. В регуляции скорости
гликогенолиза в печени большое значение имеет система циклических нуклеотидов. Кроме того, глюкоза в печени
образуется также в процессе глюконеогенеза.
Основными субстратами глюконеогенеза служат
лактат, глицерин
и аминокислоты. Принято
считать, что почти все аминокислоты,
за исключением лейцина,
могут пополнять пул предшественников
глюконеогенеза.
При оценке
углеводной функции печени необходимо иметь в виду, что соотношение между
процессами утилизации и образования глюкозы регулируется
прежде всего нейрогуморальным путем при участии желез внутренней секреции.
Центральную
роль в превращениях глюкозы
и саморегуляции углеводного обмена в печени играет глюкозо-6-фосфат. Он резко
тормозит фосфоролитическое расщепление гликогена, активирует
ферментативный перенос глюкозы
с уридиндифосфоглюкозы на молекулу
синтезирующегося гликогена,
является субстратом
для дальнейших гликолитических превращений, а также окисления глюкозы, в том числе по
пентозофосфатному пути. Наконец, расщепление глюкозо-6-фосфата фосфатазой обеспечивает
поступление в кровь свободной глюкозы, доставляемой
током крови во все органы и ткани.
Как отмечалось,
наиболее мощным аллостерическим активатором фосфофруктокиназы-1 и ингибитором
фруктозо-1,6-бисфосфатазы печени является фруктозо-2,6-бисфосфат (Ф-2,6-Р2).
Повышение в гепатоцитах уровня Ф-2,6-Р2 способствует усилению гликолиза и уменьшению
скорости глюконеогенеза.
Ф-2,6-Р2 снижает ингибирующее действие АТФ на
фосфо-фруктокиназу-1 и увеличивает сродство этого фермента к
фруктозо-6-фосфату. При ингибировании фруктозо-1,6-бисфосфатазы Ф-2,6-Р2
возрастает значение КМ для фруктозо-1,6-бисфосфата. Содержание
Ф-2,6-Р2 в печени, сердце, скелетной мускулатуре и других тканях контролируется
бифункциональным ферментом,
который осуществляет синтез Ф-2,6-Р2 из фруктозо-6-фосфата и АТФ и гидролиз его до
фруктозо-6-фосфата и Pi, т.е. фермент одновременно
обладает и киназной, и бисфосфатазной активностью.
Следует
отметить, что при генетически обусловленной нетолерантности к фруктозе или недостаточной
активности
фруктозо-1,6-бисфосфатазы наблюдается индуцируемая фруктозой гипогликемия,
возникающая вопреки наличию больших запасов гликогена. Вероятно,
фруктозо-1-фосфат и фруктозо-1,6-бисфосфат ингибируют фосфорилазу печени по аллосте-рическому
механизму.
Известно
также, что метаболизм
фруктозы по
гликолитическому пути в печени происходит гораздо быстрее, чем метаболизм глюкозы. Для метаболизма глюкозы характерна стадия,
катализируемая фосфофрукто-киназой-1. Как известно, на этой стадии
осуществляется метаболический контроль скорости катаболизма глюкозы. Фруктоза минует эту
стадию, что позволяет ей интенсифицировать в печени процессы метаболизма, ведущие к
синтезу жирных кислот, их эстерификацию и секрецию липопротеинов очень низкой
плотности; в результате может увеличиваться концентрация триглицеридов
в плазме крови.
Галактоза в печени сначала
фосфорилируется при участии АТФ и фермента галактокиназы с
образованием галактозо-1-фосфата. Для га-лактокиназы печени плода и ребенка характерны
значения КМ и Vмaкс, примерно в 5 раз превосходящие
таковые у ферментов
взрослого человека. Большая часть галактозо-1-фосфата в печени превращается в
ходе реакции, катализируемой гексозо-1-фосфат-уридилилтрансферазой:
УДФ-глюкоза +
Галактозо-1-фосфат –> УДФ-галактоза + Глюкозо-1-фосфат.
Это
уникальная трансферазная реакция возвращения галактозы в основное русло
углеводного метаболизма.
Наследственная утрата
гексозо-1-фосфат-уридилилтрансферазы приводит к галактоземии – заболеванию, для
которого характерны умственная отсталость и катаракта хрусталика. В этом случае
печень новорожденных теряет способность метаболизи-ровать D-галактозу, входящую
в состав лактозы
молока.
РОЛЬ ПЕЧЕНИ В ЛИПИДНОМ ОБМЕНЕ
Ферментные
системы печени способны катализировать все реакции или значительное большинство
реакций метаболизма
липидов. Совокупность этих
реакций лежит в основе таких процессов, как синтез высших жирных кислот,
триглицеридов, фосфолипидов,
холестерина и его эфиров,
а также липолиз триглицеридов, окисление жирных кислот,
образование ацетоновых (кетоновых) тел и т.д. Напомним, что ферментативные
реакции синтеза триглицеридов в печени и жировой ткани сходны. Так,
КоА-производные жирной кислоты с длинной цепью взаимодействуют с
глицерол-3-фосфатом с образованием фосфатидной кислоты, которая затем
гидролизуется до диглицерида. Путем присоединения к последнему еще одной молекулы КоА-производного
жирной кислоты образуется триглицерид. Синтезированные в печени три-глицериды
либо остаются в печени, либо секретируются в кровь в форме липопротеинов. Секреция происходит с
известной задержкой (у человека 1–3 ч). Задержка секреции, вероятно, соответствует
времени, необходимому для образования липопротеинов. Основным
местом образования плазменных пре-β-липопротеинов (липопротеины очень
низкой плотности – ЛПОНП) и α-липопротеинов (липопротеины высокой
плотности – ЛПВП) является печень.
РОЛЬ ПЕЧЕНИ В ОБМЕНЕ БЕЛКОВ
Печень играет
центральную роль в обмене белков.
Она выполняет следующие основные функции: синтез специфических белков плазмы; образование мочевины и мочевой кислоты; синтез холина и креатина; трансаминирование и дезаминирование аминокислот, что весьма
важно для взаимных превращений аминокислот, а также для
процесса глюконеогенеза
и образования кетоновых тел. Все альбумины плазмы, 75–90%
α-глобу-линов и 50% β-глобулинов синтезируются гепатоцитами. Лишь
γ-гло-булины продуцируются не гепатоцитами, а системой макрофагов, к
которой относятся звездчатые ретикулоэндотелиоциты (клетки Купфера). В основном
γ-глобулины образуются в печени. Печень является единственным органом, где
синтезируются такие важные для организма белки, как протромбин, фибриноген, проконвертин и
проакцелерин.
При
заболеваниях печени определение фракционного состава белков плазмы (или сыворотки)
крови нередко представляет интерес как в диагностическом, так и в
прогностическом плане. Известно, что патологический процесс в гепатоцитах резко
снижает их синтетические возможности. В результате содержание альбумина в плазме крови резко падает,
что может привести к снижению онкотического давления плазмы крови, развитию
отеков, а затем асцита. Отмечено, что при циррозах печени, протекающих с
явлениями асцита, содержание альбуминов
в сыворотке крови
на 20% ниже, чем при циррозах без асцита.
Нарушение
синтеза ряда белковых факторов системы свертывания крови
при тяжелых заболеваниях печени может привести
к геморрагическим явлениям.
При
поражениях печени нарушается также процесс дезаминирования
аминокислот,
что способствует увеличению их концентрации
в крови и моче. Так, если в норме содержание азота
аминокислот
в сыворотке
крови составляет примерно 2,9–4,3 ммоль/л, то при тяжелых заболеваниях
печени (атрофические процессы) эта величина возрастает до 21 ммоль/л, что
приводит к аминоацидурии. Например, при острой атрофии печени количество тирозина
в суточном количестве мочи может достигать 2 г (при норме 0,02–0,05 г/сут).
В
организме
образование мочевины
в основном происходит в печени. Синтез мочевины
связан с затратой довольно значительного количества энергии (на образование 1 молекулы
мочевины
расходуется 3 молекулы
АТФ). При заболевании печени, когда количество АТФ
в гепатоцитах уменьшено, синтез мочевины
нарушается. Показательно в этих случаях определение в сыворотке отношения азота
мочевины
к аминоазоту. В норме это отношение равно 2:1, а при тяжелом поражении печени
составляет 1:1.
Большая
часть мочевой
кислоты также образуется в печени, где много фермента
ксантиноксидазы, при участии которого оксипурины (гипо-ксантин и ксантин)
превращаются в мочевую
кислоту. Нельзя забывать о роли печени и в синтезе креатина. Имеются два
источника креатина в организме.
Существует экзогенный креатин, т.е. креатин пищевых продуктов (мясо, печень и
др.), и эндогенный креатин, синтезирующийся в тканях.
Синтез креатина происходит в основном в печени, откуда он с током крови
поступает в мышечную ткань.
Здесь креатин, фосфори-лируясь, превращается в креатинфосфат, а из последнего
образуется креатин.
Чужеродные
вещества
(ксенобиотики) в печени нередко превращаются в менее токсичные и даже
индифферентные вещества.
По-видимому, только в этом смысле можно говорить об «обезвреживании» их в
печени. Происходит это путем окисления,
восстановления,
метилирования,
ацетилирования
и конъюгации с теми или иными веществами.
Необходимо отметить, что в печени окисление,
восстановление
и гидролиз
чужеродных соединений осуществляют в основном микросомальные ферменты.
Наряду с микро-сомальным в печени существует также пероксисомальное окисление.
Пероксисомы – микротельца, обнаруженные в гепатоцитах; их можно рассматривать
как специализированные окислительные органеллы. Эти микротельца содержат
оксидазу мочевой
кислоты, лактатоксидазу, окси-дазу D-аминокислот, а также каталазу.
Последняя катализирует расщепление перекиси водорода,
которая образуется при действии указанных оксидаз; отсюда и название этих
микротелец – пероксисомы. Пероксисо-мальное окисление,
так же как и микросомальное, не сопровождается образованием макроэргических
связей.
В
печени широко представлены также «защитные» синтезы, например синтез мочевины,
в результате которого обезвреживается весьма токсичный аммиак.
В результате гнилостных процессов, протекающих в кишечнике, из тирозина
образуются фенол
и крезол,
а из триптофона – скатол
и индол.
Эти вещества
всасываются и с током крови поступают в печень, где обезвреживаются путем
образования парных соединений с серной или глюкуроновой
кислотой.
Обезвреживание
фенола,
крезола,
скатола
и индола
в печени происходит в результате взаимодействия этих соединений не со
свободными серной и глюкуроновой
кислотами, а с их так называемыми активными формами: ФАФС и УДФГК.
Глюкуроновая
кислота участвует не только в обезвреживании продуктов гниения белковых веществ,
образовавшихся в кишечнике, но и в связывании ряда других токсичных соединений,
образующихся в процессе обмена в тканях.
В частности, свободный, или непрямой, билирубин,
обладающий значительной токсичностью,
в печени взаимодействует с глюкуроновой
кислотой, образуя моно- и диглюкурониды билирубина.
Нормальным метаболитом
является и гиппуровая кислота, образующаяся в печени из бензойной
кислоты и глицина.
Синтез
гиппуровой кислоты у человека протекает преимущественно в печени. Поэтому в
клинической практике довольно часто для выяснения антитоксической функции
печени применяют пробу Квика–Пытеля (при нормальной функциональной способности
почек): после нагрузки бензо-атом натрия
в моче определяют количество образовавшейся гиппуровой кислоты. При
паренхиматозных поражениях печени синтез гиппуровой кислоты снижен.
В
печени широко представлены процессы метилирования.
Так, перед выделением с мочой амид никотиновой
кислоты (витамин РР) метилируется в печени; в результате образуется
N-метилникотинамид. Наряду с метилированием
интенсивно протекают и процессы ацетилирования
. В частности, в печени ацетилированию
подвергаются различные сульфаниламидные
препараты.
Примером
обезвреживания токсичных продуктов в печени путем восстановления
является превращение нитробензола
в парааминофенол. Многие ароматические углеводы
обезвреживаются путем окисления
с образованием соответствующих карбоновых
кислот.
Печень
принимает активное участие в инактивации различных гормонов.
С током крови гормоны
попадают в печень, при этом активность
их в большинстве случаев резко снижается или полностью утрачивается. Так, стероидные
гормоны, подвергаясь микросомальному окислению,
инакти-вируются, превращаясь затем в соответствующие глюкурониды и сульфаты.
Под влиянием аминооксидаз в печени происходит окисление
ка-техоламинов и т.д.
Из
приведенных примеров видно, что печень способна инактивировать ряд
сильнодействующих физиологических и чужеродных (в том числе токсичных) веществ.
Рассмотрим
только гемохромогенные пигменты,
которые образуются в организме
при распаде гемоглобина
(в значительно меньшей степени при распаде миоглобина,
цитохромов
и др.). Распад гемоглобина
протекает в клетках
макрофагов, в частности в звездчатых ретикулоэндотелиоцитах, а также в
гистиоцитах соединительной ткани
любого органа.
Начальным
этапом распада гемоглобина
является разрыв одного метинового мостика с образованием вердоглобина. В
дальнейшем от молекулы
вердоглобина отщепляются атом
железа
и белок
глобин.
В результате образуется биливердин, который представляет собой цепочку из
четырех пиррольных колец, связанных метановыми мостиками. Затем биливердин,
восстанавливаясь, превращается в билирубин
– пигмент,
выделяемый с желчью и поэтому называемый желчным
пигментом. Образовавшийся билирубин
называется непрямым (неконъю-гированным) билирубином.
Он нерастворим в воде,
дает непрямую реакцию с диазореактивом, т.е. реакция протекает только после
предварительной обработки спиртом.
В
печени билирубин
соединяется (конъюгирует) с глюкуроновой
кислотой. Эта реакция катализируется ферментом
УДФ-глюкуронилтранс-феразой, при этом глюкуроновая
кислота вступает в реакцию в активной форме, т.е. в виде УДФГК.
Образующийся глюкуронид билирубина
получил название прямого билирубина
(конъюгированный билирубин). Он растворим в воде
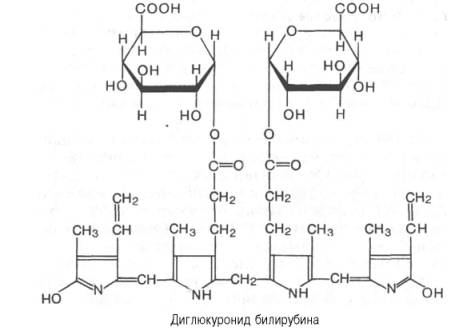
и дает прямую реакцию с диазореактивом. Большая часть билирубина
соединяется с двумя молекулами
глюкуроновой
кислоты, образуя диглюкуронид билирубина:
Образовавшийся
в печени прямой билирубин
вместе с очень небольшой частью непрямого билирубина
выводится с желчью в тонкую кишку. Здесь от прямого билирубина
отщепляется глюкуроновая
кислота и происходит его восстановление
с последовательным образованием мезобилирубина и мезобилиногена
(уробилиногена). Принято считать, что около 10% билирубина
восстанавливается до мезобилиногена на пути в тонкую кишку, т.е. во
внепеченочных желчных путях и в желчном пузыре. Из тонкой кишки часть
образовавшегося мезобилиногена (уробилиногена) резорбируется через кишечную
стенку, попадает в воротную вену и током крови переносится в печень, где
расщепляется полностью до ди- и трипирролов. Таким образом, в норме в общий
круг кровообращения и мочу мезобилиноген не попадает.
Основное
количество мезобилиногена из тонкой кишки поступает в толстую и здесь
восстанавливается до стеркобилиногена при участии анаэробной микрофлоры.
Образовавшийся стеркобилиноген в нижних отделах толстой кишки (в основном в прямой
кишке) окисляется до стерко-билина и выделяется с калом. Лишь небольшая часть
стеркобилиногена всасывается в систему нижней полой вены (попадает сначала в
геморроидальные вены) и в дальнейшем выводится с мочой. Следовательно, в норме
моча человека содержит следы стеркобилиногена (за сутки его выделяется с мочой
до 4 мг). К сожалению, до последнего времени в клинической практике
стеркобилиноген, содержащийся в нормальной моче, продолжают называть
уробилиногеном. На рис. схематично показаны пути образования уробилиногеновых
тел в организме
человека. В клинической практике укоренился термин «уробилиноген мочи». Под
этим термином следует понимать те производные билирубина
(билирубиноиды), которые обнаруживаются в моче. Положительная реакция на
уробилиноген может быть обусловлена повышенным содержанием того или иного
билирубиноида в моче и является, как правило, отражением патологии.
Определение
в клинике содержания билирубина
в крови (общего, непрямого и прямого), а также уробилиногена мочи имеет важное
значение при дифференциальной диагностике желтух различной этиологии. При
гемолитической желтухе («надпеченочной») вследствие повышенного гемолиза
эритроцитов
и разрушения гемоглобина
происходит интенсивное образование непрямого билирубина
в ретикулоэндотелиальной системе, б). Печень оказывается неспособной
утилизировать такое большое количество непрямого билирубина,
что приводит к его накоплению в крови и тканях.
В печени при этом синтезируется повышенное количество прямого билирубина,
который с желчью попадает в кишечник. В тонкой кишке в повышенных количествах
образуется мезобилиноген и в последующем – стеркобилиноген. Всосавшаяся часть
мезобилиногена утилизируется печенью, а резорбирующийся в толстой кишке
стеркобилиноген выводится с мочой. Таким образом, для гемолитической желтухи в
типичных случаях характерны следующие клинико-лабораторные показатели:
повышение уровня общего и непрямого билирубина
в крови, в моче – отсутствие билирубина
(непрямой билирубин
не фильтруется почками) и положительная реакция на уробилиноген (за счет
повышенного попадания в кровь и мочу стеркобилиногена, а в тяжелых случаях – и
за счет мезобилиногена, не утилизирующегося печенью); лимонно-желтый оттенок
кожных покровов (сочетание желтухи и анемии); увеличение размеров селезенки;
ярко окрашенный кал.
При
механической (обтурационной, или «подпеченочной») желтухе, в) нарушен отток
желчи (закупорка общего желчного протока камнем, рак
головки поджелудочной железы). Это приводит к деструктивным изменениям в печени
и попаданию элементов желчи (билирубин, холестерин,
желчные кислоты) в кровь. При полной обтурации общего желчного протока желчь не
попадает в кишечник, поэтому образования в кишечнике билирубиноидов не
происходит, кал обесцвечен и реакция на уробилиноген мочи отрицательная. Таким
образом, при механической желтухе в крови повышено количество общего билирубина
(за счет прямого), увеличено содержание холестерина
и желчных
кислот, а в моче – высокий уровень билирубина
(прямого). Клиническими особенностями обтурационной желтухи являются яркая
желтушная окраска кожи,
бесцветный кал, зуд кожи
(раздражение нервных окончаний желчными
кислотами, отлагающимися в коже). Следует заметить, что при длительно
сохраняющейся механической желтухе могут существенно нарушаться функции печени,
в том числе одна из главных – детоксикационная. В этом случае может произойти
частичный «отказ» печени от непрямого билирубина,
что может привести к его накоплению в крови. Иными словами, увеличение уровня
фракции непрямого билирубина
при механической желтухе является плохим прогностическим признаком.
При
паренхиматозной («печеночной») желтухе, возникающей чаще всего при ее вирусном
поражении, развиваются воспалительно-деструктивные процессы в печени, ведущие к
нарушению ее функций. На начальных этапах гепатита процесс захвата и
глюкуронирования непрямого билирубина
сохраняется, однако образующийся прямой билирубин
в условиях деструкции печеночной паренхимы
частично попадает в большой круг кровообращения, что ведет к желтухе. Экскреция
желчи также нарушена, билирубина
в кишечник попадает меньше, чем в норме. Меньше обычного образуется
мезобилиногена, и меньшее количество его всасывается в кишечнике. Однако даже
это небольшое количество поступающего в печень мезобилиногена не усваивается
ею. Мезобилиноген, «уклоняясь», попадает в кровь, а затем выделяется с мочой,
что предопределяет положительную реакцию на уробилиноген. Количество
образующегося стеркобилиногена также снижено, поэтому кал гипохоличный. Итак,
при паренхиматозной желтухе отмечается повышение в крови концентрации
общего билирубина,
преимущественно за счет прямого. В кале снижено содержание стеркобилиногена.
Реакция на уробилиноген мочи положительная за счет попадания в мочу
мезобилиногена. Следует отметить, что при прогрессирующем гепатите, когда
печень утрачивает свою детоксикационную функцию, в крови накапливается
значительное количество и непрямого билирубина.
Кроме того, при резко выраженном воспалении печени, ее «набухании», может
произойти сдавление желчных капилляров и протоков, возникнуть внутрипеченочный
холестаз, что придает паренхиматозной желтухе черты механической с
соответствующей клинико-лабораторной картиной (ахоличный кал, отсутствие
реакции на уробилиноген).
В
табл. приведены наиболее характерные сдвиги клинико-лабо-раторных показателей
при различных типах желтух.
Следует
иметь в виду, что в практике редко наблюдается желтуха какого-либо одного типа
в «чистом» виде. Чаще встречается сочетание того или иного типа. Так, при
выраженном гемолизе
неизбежно страдают различные органы, в том числе и печень, что может привнести
элементы паренхиматозной желтухи при гемолизе.
В свою очередь паренхиматозная желтуха, как правило, включает в себя элементы
механической. При механической желтухе, возникающей вследствие сдавливания
большого сосочка двенадцатиперстной кишки (фатерова соска) при раке
головки поджелудочной железы, неизбежен повышенный гемолиз
как следствие раковой интоксикации.
Желчь
– жидкий секрет желтовато-коричневого цвета, отделяется печеночными клетками.
В сутки у человека образуется 500–700 мл желчи (10 мл на 1 кг массы тела).
Желчеобразование происходит непрерывно, хотя интенсивность этого процесса на
протяжении суток резко колеблется. Вне пищеварения печеночная желчь переходит в
желчный пузырь, где происходит ее сгущение
в результате всасывания воды
и электролитов.
Относительная плотность печеночной желчи 1,01, а пузырной – 1,04. Концентрация
основных компонентов в пузырной желчи в 5–10 раз выше, чем в печеночной.
Предполагают,
что образование желчи начинается с активной секреции
гепатоцитами воды,
желчных
кислот и билирубина,
в результате которой в желчных канальцах появляется так называемая первичная
желчь. Последняя, проходя по желчным ходам, вступает в контакт с плазмой
крови, вследствие чего между желчью и плазмой
устанавливается равновесие электролитов,
т.е. в образовании желчи принимают участие в основном два механизма –
фильтрация и секреция.
В
печеночной желчи можно выделить две группы веществ.
Первая группа – это вещества,
которые присутствуют в желчи в количествах, мало отличающихся от их концентрации
в плазме
крови (например, ионы
Na+, К+, креатин и др.), что в какой-то мере служит
доказательством наличия фильтрационного механизма. Ко второй группе относятся
соединения, концентрация
которых в печеночной желчи во много раз превышает их содержание в плазме
крови (билирубин, желчные
кислоты и др.), что свидетельствует о наличии секреторного механизма. В
последнее время появляется все больше данных о преимущественной роли активной секреции
в механизме желчеобразования. Кроме того, в желчи обнаружен ряд ферментов,
из которых особо следует отметить щелочную фосфатазу
печеночного происхождения. При нарушении оттока желчи активность
данного фермента
в сыворотке
крови возрастает.
Основные
функции желчи. Эмульсификация. Соли желчных
кислот обладают способностью
значительно уменьшать поверхностное
натяжение. Благодаря этому они
осуществляют эмульгирование жиров в кишечнике, растворяют жирные кислоты и нерастворимые
в воде мыла. Нейтрализация кислоты. Желчь, рН которой немногим
более 7,0, нейтрализует кислый химус, поступающий из желудка, подготавливая его
для переваривания в кишечнике. Экскреция. Желчь – важный носитель экскрети-руемых желчных
кислот и холестерина. Кроме того, она удаляет из организма многие лекарственные вещества, токсины, желчные
пигменты и различные
неорганические вещества, такие, как медь, цинк и ртуть. Растворение холестерина. Как отмечалось, холестерин, подобно высшим
жирным кислотам, представляет
собой нерастворимое в воде соединение, которое сохраняется в желчи в
растворенном состоянии лишь благодаря присутствию в ней солей желчных
кислот и фосфатидилхолина. При недостатке желчных
кислот холестерин выпадает в осадок, при этом могут образовываться
камни. Обычно камни имеют окрашенное желчным
пигментом внутреннее ядро, состоящее
из белка. Чаще всего встречаются камни, у которых ядро
окружено чередующимися слоями холестерина и билирубината кальция. Такие камни содержат до 80% холестерина. Интенсивное образование камней отмечается при
застое желчи и наличии инфекции. При застое желчи встречаются камни, содержащие
90–95% холестерина, а при инфекции могут образовываться камни,
состоящие из билирубината кальция. Принято считать, что присутствие бактерий
сопровождается увеличением β-глюкуронидазной активности желчи, что приводит к расщеплению конъюгатов билирубина; освобождающийся билирубин служит субстратом для образования камней.


{kind=link}