

МОЛЕКУЛЯРНО-КЛІТИННІ МЕХАНІЗМИ ДІЇ ГОРМОНІВ на клітини-мішені. Роль гормонів в регуляції метаболічних процесів. Вітаміни як компоненти харчування. Водорозчинні та жиророзчинні вітаміни
Загальні відомості про гормони
Гормони – це біологічно активні речовини різноманітної хімічної природи, які виробляються в спеціалізованих клітинах залоз внутрішньої секреції, надходять безпосередньо в кров і здійснюють гуморальну регуляцію обміну речовин і функцій організму.
Концентрація гормонів в крові дуже низька, від мікромолярної (10‑6 моль/л) до пікомолярної (10-12 моль/л), але кількість молекул, яка відповідає цій концентрації, величезна – 1011-1017 молекул/л, практично трильйони молекул у 1 літрі крові. Ця величезна кількість молекул гормонів робить можливим їх вплив на кожну окрему клітину організму. Але гормони діють не на всі клітини, а лише на клітини-мішені, що містять специфічні білки-рецептори, які зв'язують молекули гормонів із високою вибірковістю. Рецептори локалізовані у плазматичній мембрані клітин або їх цитоплазмі чи ядрі. Кількість рецепторів у клітині не постійна і регулюється або кількістю власного гормону, або дією іншого гормону. На плазматичній мембрані кількість рецепторів може досягати десятків тисяч.
Залози внутрішньої секреції
За́лози вну́трішньої секреції або ендокринні залози — це ті, які не мають вивідних проток, і виділяють свою «продукцію» — гормони, безпосередньо в кров, яка омиває ці залози. Саме через це їх називають залозами внутрішньої секреції (ендокринні залози).
У людини до ендокринних залоз відносять: епіфіз, гіпофіз, щитоподібну, загрудинну, паращитоподібні, наднирники, острівці Лангерганса підшлункової залози та ендокринну частину статевих залоз (рис. 1). У зв'язку з цим підшлункову залозу та статеві залози відносять до змішаних залоз за типом секреції.

Рис 1. Основні залози внутрішньої секреції
Більшість гормональних процесів регулюється гіпофізом, який виділяє тропні гормони, що регулюють діяльність інших залоз. В свою чергу, секреторну діяльність гіпофіза (рис. 2) регулює гіпоталамус, нейрони якого продукують нейроендокринні трансмітери, що стимулюють або пригнічують роботу гіпофіза. Ці два органи утворюють єдину гіпоталамо-гіпофізарну систему, в якій перший виконує регулюючу роль, а інший — ефекторну.
Рис. 2. Гіпофіз
Кінцеві ефекти дії гормонів:
1. Зміна мембранної проникності. Змінюючи стан мембран, гормони можуть посилювати чи гальмувати швидкість переходу ферментів, коферментів чи субстратів в клітину і з клітини. В результаті може змінюватись швидкість і напрямок ферментативних процесів.
2. Зміна активності окремих ферментів. Ферменти мають алостеричні центри, а гормони можуть бути алостеричними ефекторами. Діючи на алостеричні центри, вони можуть змінити конформац3ію фермента, в тому чилі активного центру, тим самим пригнічуючи або активуючи дію ферменту. Зрозуміло, що так діяти можуть лише гормони, які проникають в клітину.
3. Зміна інтенсивності синтезу фермента, тобто, гормони можуть впливати на генетичний апарат клітини. Так діють кортикостероїди, андрогени, естрогени. Вони взаємодіють із специфічними рецепторами в цитоплазмі клітини. Комплекс гормон-рецептор переходить в ядро, де зв»язується з негістоновими білками хроматину. Ці білки можуть набувати значного негативного заряду і витісняти з хроматину пістони. Ділянка ДНК звільняється (оперон), в цих місцях починається транскрипція і синтез певного ферменту.
4. Вплив на розпад ферментів, синтез коферментів.
Класифікація гормонів
Існують морфологічна, хімічна, фізіологічна класифікації гормонів. За морфологічною класифікацією гормони розділяють залежно від місця їх синтезу, наприклад, гормони гіпофіза, щитовидної залози, підшлункової залози, надниркових залоз, статевих залоз тощо. Але ряд фактів не відповідають такому поділу. Так, статеві гормони утворюються в різних місцях: статевих залозах, корі надниркових залоз.
Деякі гормони гіпоталамуса наявні в інших відділах мозку, шишкоподібній залозі, шлунково-кишковому тракті. А головним є те, що не тільки в ендокринних залозах, а майже у всіх органах і тканинах організму є клітини, в яких синтезуються гормони. Ці клітини різних органів об'єднуються в АПУД-систему і називаються апудоцитами. Апудоцити виробляють катехоламіни, гістамін, серотонін, мелатонін, деякі гормони гіпофіза, гастрин, секретин тощо.
За хімічною природою гормони поділяються на такі групи (табл. 1):
1) Білково-пептидні гормони
2) Гормони — похідні амінокислот
3) Гормони стероїдної природи
4) Біорегулятори — похідні арахідонової кислоти
За біологічними функціями гормони ділять на такі групи:
1. Гормони, що регулюють обмін вуглеводів, жирів, амінокислот: інсулін, глюкагон, глюкокортикоїди, адреналін.
2. Гормони, що регулюють водно-сольовий обмін: альдостерон, вазопресин, ангіотензин, натрійуретичний фактор передсердя.
3. Гормони, що регулюють обмін кальцію і фосфатів: паратгормон, кальцитонін, активні форми вітаміну D.
4. Гормони, що відповідають за репродуктивну функцію організму: андрогени, естрогени, прогестерон, гонадотропні гормони, пролактин.
5. Гормони, що регулюють функції периферичних ендокринних залоз: гормони гіпоталамуса, тропні гормони гіпофіза.
У цій класифікації не враховується поліфункціональність ряду гормонів. Наприклад, адреналін регулює не тільки обмін речовин, але й частоту серцевих скорочень, кров'яний тиск, зменшує спазм бронхів. Деякі гормони не включені в класифікацію за функціями, оскільки викликають різні зміни, серед яких не виділено первинних (тироксин, соматотропін).
Регуляція синтезу і секреції гормонів
Ендокринні залози є складовою частиною системи нейрогуморальної регуляції організму. Під впливом різноманітних зовнішніх і внутрішніх подразників виникають електричні імпульси (потенціали дії) у спеціалізованих дуже чутливих рецепторах, що передаються доцентровими нервовими волокнами до клітин ЦНС. Після обробки інформації в ЦНС сигнали передаються на периферію (рис. 3). Під прямим контролем нервової системи знаходяться гіпоталамус і мозкова речовина надниркових залоз. Інші ендокринні залози зв'язані з нервовою системою опосередковано через гормони гіпоталамуса і гіпофіза.
Рис. 3. Зв’язок ендокринної та нервової систем
(суцільні стрілки позначають синтез гормону, пунктирні – вплив гормону на органи мішені)
У відповідь на сигнали із ЦНС гіпоталамус синтезує і секретує гіпоталамічні регуляторні гормони двох типів – ліберини і статини, які через систему портального кровообігу гіпофіза надходять до клітин аденогіпофіза. Кожний гіпоталамічний гормон регулює секрецію якогось одного гормону передньої частки гіпофіза. Ліберини стимулюють секрецію гормону гіпофіза, а статини пригнічують. Гормони аденогіпофіза, які називаються тропними або тропінами, виділяються в кров, транспортуються до певної ендокринної залози, стимулюють утворення і секрецію нею гормонів. Гормони периферичних залоз діють на органи і тканини-мішені, викликаючи відповідні фізіологічні й біологічні зміни. Із точки зору переносу інформації багатоступеневий процес можна розглядати як "посилення потоку інформації".
Синтез і секреція гормонів всіх видів регулюються механізмами, що працюють за принципом позитивного і негативного зворотних зв'язків. Так, концентрація у крові гормонів периферичних залоз чи тропних гормонів гіпофіза впливає на секрецію гормонів гіпоталамуса і гіпофіза.
Наприклад, підвищений вміст у крові тироксину гальмує секрецію тиреоліберину гіпоталамусом і тиреотропіну гіпофізом. На швидкість секреції гормонів ендокринними залозами впливають також наявні у крові продукти метаболізму, іони. Секреція деяких гормонів підпорядковується певним біологічним ритмам. Таким чином, як тільки гормон починає діяти на чутливу до нього клітину чи групу клітин, одночасно виникає сигнал, котрий гальмує дію гормону. Цим сигналом є або підвищений вміст іншого гормону, або корекція показника гомеостазу, зміна якого була первинною причиною активації певної залози.
У результаті надлишкового чи недостатнього утворення гормонів розвиваються ендокринні захворювання. Підвищення продукції гормонів може бути наслідком злоякісного перетворення клітин ендокринної залози. Зниження продукції гормонів зв'язане з незворотними пошкодженнями чи загибеллю клітин залози. Причиною порушень регуляції і синтезу гормонів є генетичні дефекти клітин ендокринної залози або білків рецепторів клітин-мішеней, але звичайно має місце не дефект якогось одного гена, як при більшості молекулярних хвороб, а вроджені порушення ряду генів, тобто спостерігається багатофакторна спадкова схильність до розвитку патологічного процесу.
Для біохімічної діагностики ендокринних захворювань визначають в крові та інших біологічних рідинах концентрацію гормонів, продуктів їх обміну, а також вміст метаболітів, іонів, що регулюється відповідними гормонами.
Механізм дії гормонів
Механізм дії гормонів залежить від здатності їх проникати через плазматичну мембрану клітини. Водорозчинні гормони білково-пептидної природи, а також адреналін не проходять через плазматичну мембрану, а взаємодіють із специфічними мембранними рецепторами. Внаслідок взаємодії включаються внутрішньоклітинні шляхи передачі інформації, які регулюють метаболізм клітини та різноманітні клітинні процеси. На рівні плазматичної мембрани передача інформації здійснюється шляхом послідовної зміни конформації мембранних білків (рецепторного, сполучного) і ферменту. Останній розміщений із внутрішньої сторони мембрани і каталізує утворення низькомолекулярної речовини – вторинного посередника, месенджера. Дифузія вторинного посередника забезпечує швидке поширення сигналу по всій клітині до конкретних ферментів чи інших білків, які реалізують відповідь клітини на первинний сигнал – гормон чи іншу речовину (наприклад, ліки, бактеріальний токсин), що здатні зв'язуватись із гормональним рецептором плазматичної мембрани.
Безпосередньою мішенню дії вторинних посередників служать ферменти протеїнкінази, які шляхом фосфорилювання активують чи інгібують специфічні клітинні білки. Усі складові компоненти складають систему (каскад) і забезпечують ефективну передачу і підсилення відповідного гормонального сигналу.
Першою була відкрита аденілатциклазна месенджерна система, в якій вторинним посередником є циклічний АМФ. Структура цАМФ, реакція синтезу, яку каталізує мембранозв'язана аденілатциклаза, і реакція розпаду під дією фосфодіестерази розглянуті раніше. Сигнал з гормональних рецепторів на аденілатциклазу передають G-білки двох типів: Gs‑білок активує аденілатциклазу, а Gi-білок гальмує. G-білки обох типів складаються з альфа-, бета- і гама-субодиниць, причому відрізняються альфа-субодиницями, мають центри зв'язування ГТФ і ГДФ та здатні гідролізувати зв'язаний ГТФ до ГДФ і неорганічного фосфату. До включення системи G-білок містить зв'язаний з альфа-субодиницею ГДФ і не взаємодіє з аденілатциклазою.
Приєднання гормону зумовлює конформаційні зміни рецептора і G‑білка. Останній швидко зв'язує ГТФ замість ГДФ і в такій формі змінює активність аденілатциклази (активує чи гальмує, залежно від типу). Одночасно стимулюється ГТФазна активність G-білка і після переходу ГТФ у ГДФ активація аденілатциклази припиняється. Білок-рецептор, G-білок і каталітична субодиниця аденілатциклази разом складають аденілатциклазний комплекс. Трансмембранна передача сигналу комплексом завершується утворенням цАМФ. Далі передача сигналу пов'язана з дією цАМФ на внутрішньоклітинні компоненти аденілатциклазної системи (рис. 4).
Рис. 4. Схема аденілатциклазної системи
Робота аденілатциклазної системи детально описуєтьcя при розгляді регуляції обміну глікогену, де адреналін запускає каскадний процес активації глікогенфосфорилази та інактивації глікогенсинтетази. Багатостадійність системи має важливе значення, оскільки в такому каскадному процесі початковий гормональний сигнал зазнає багатократного підсилення.
При дії на клітини-мішені соматостатину, ангіотензину II, нейрогормонів енкефалінів і ендорфінів, а також при зв'язуванні катехоламінів з альфа2-рецепторами сигнал від рецептора передається через Gі-білки, що зумовлює гальмування активності аденілатциклази, зниження рівня в клітині цАМФ і активності відповідних протеїнкіназ.
На аденілатциклазний шлях передачі сигналу в клітину впливають деякі бактеріальні екзотоксини. Наприклад, токсин, який утворює холерний вібріон, каталізує реакцію переносу АДФ-рибози з НАД на альфа-ланцюг Gs-білка (реакція рибозилювання), що переводить білок у постійно активний стан з втратою чутливості до гормональних сигналів. Внаслідок цього в клітинах кишечника зростає рівень цАМФ, який викликає інтенсивну секрецію кишкового соку. Екзотоксин коклюшу каталізує рибозилювання альфа-субодиниці Gi-білка, що зумовлює блокаду передачі гальмівних сигналів від гормональних рецепторів на аденілатциклазу. В результаті активність аденілатциклази і рівень цАМФ в клітинах також зростають.
Друга система передачі гормональних сигналів – фосфоінозитидна – складніша за аденілатциклазну. В ній використовується комбінація трьох вторинних посередників – інозитолтрифосфату, діацилгліцеролу і іонів Са2+.
Перші дві сполуки утворюються при гідролізі мембранного фосфоліпіду фосфатидилінозитол-4,5-дифосфату. Реакцію каталізує фосфоліпаза С, яка переходить в активний стан у результаті приєднання гормону до рецептора. Сигнал від рецептора до фосфоліпази С також передають G-білки.
Через фосфоінозитидну систему реалізується дія катехоламінів (при їх зв'язуванні з альфа1-адренорецепторами), тироліберину, гонадоліберину, вазопресину, ангіотензину II, гастрину, холецистокініну, брадикініну та інших гормонів.
Вторинні посередники фосфоінозитидної системи діють синергічно на клітинні функції, але механізм їх дії різний. Водорозчинний інозитолтрифосфат (IФ3) дифундує у цитозоль, зв'язується з рецепторами мембрани ендоплазматичного ретикулума і зумовлює вихід іонів Са2+ через кальцієві канали. В результаті в цитоплазмі швидко зростає рівень іонів Са2+, які зв'язуються з специфічними внутрішньоклітинними білками і активують Са2+-залежні протеїнкінази (рис. 5).
Іони Са2+ можуть бути внутрішньоклітинним посередником передачі інформації і без активації фосфоінозитидної системи, коли концентрація їх у цитоплазмі зростає внаслідок надходження ззовні через кальцієві канали плазматичної мембрани, які відкриваються в результаті зміни трансмембранного потенціалу чи під впливом певних регуляторних молекул.
Рис. 5. Схема Са2+- фосфоінозитидної системи
Г – гормон, R – рецептор Gp – ГТФ – зв’язуючий білок, який передає сигнал на ферменти, ФЛ-С – фосфоліпаза С, ФЛ-А2, ФІФ2 – фосфатидил-4,5-дифосфат, ІФ3 – інозитолтрифосфат, ДАГ - діацилгліцерол
Проте функціональна активність іонних каналів залежить від протеїнкіназного фосфорилювання білків-компонентів каналів.
Інший вторинний посередник фосфоінозитидної системи – діацилгліцерол – переводить в активний стан мембранозв'язану протеїнкіназу С, яка фосфорилює білки, специфічні для кожного типу клітин. Активність протеїнкінази С додатково стимулюють іони Са2+ і фосфатидилсерин – компонент мембран. Спільна дія Са2+- залежних протеїнкіназ і протеїнкінази С зумовлює активацію шляхом фосфорилювання ряду клітинних білків, які беруть участь у реалізації повноцінної клітинної відповіді (секреції гормонів, нейромедіаторів, ферментів, скорочення м'язів, агрегації тромбоцитів, регуляції процесів метаболізму, транспорту іонів, глюкози та інших речовин через мембрани).
Припинення передачі гормонального сигналу через фосфоінозитидну систему здійснюється завдяки інактивації вторинних посередників і дефосфорилюванню фосфорильованих білків. Від інозитолтрифосфату (ІФ3) поступово шляхом гідролізу відщеплюються фосфатні групи:
![]()
Перетворення інозитолмонофосфату у вільний інозитол під дією ферменту інозитолмонофосфатази інгібують іони літію. Тому надходження літію в організм призводить до порушень обміну фосфоінозитидів і послаблення залежних від них процесів. Цей ефект іонів Li+ лежить, вірогідно, в основі терапевтичної дії їх при маніакально-депресивних психозах.
Діацилгліцерол інактивується двома шляхами. Частина його перетворюється у фосфатидну кислоту, а інша розщеплюється до вихідних компонентів – гліцерину і жирних кислот, зокрема арахідонової – попередника простагландинів і лейкотрієнів. Із фосфатидної кислоти і інозитолу синтезується фосфатидилінозитол і далі фосфатидилінозитол-4,5-дифосфат. Для цього використовуються молекули ЦТФ і АТФ. На схемі показано цикл обміну інозитолфосфоліпідів, який об'єднує утворення вторинних посередників для передачі зовнішних сигналів, їх інактивацію і ресинтез вихідного субстрату.
Передача сигналів через фосфоінозитидну систему супроводжується зростанням концентрації в клітині ще одного вторинного посередника – циклічного ГМФ. Синтезується цГМФ під дією гуанілатциклази. Активують гуанілатциклазу арахідонова кислота, яка вивільняється при розщепленні діацилгліцеролу і фосфоліпідів мембран, та продукти її перетворення – простагландини і лейкотрієни. цГМФ активує протеїнкіназу G, що, як і протеїнкінази А, С і Са2+-залежні, фосфорилює ряд клітинних білків.
Під дією цГМФ гальмується процес передачі сигналу через фосфоінозитидну систему, що забезпечує негативний зворотний зв'язок у ній. У більшості клітин аналогічно пригнічує проведення сигналу через фосфоінозитидну систему підвищений рівень цАМФ, тобто активація аденілатциклазної системи. Таким чином попереджується надмірна інтенсивність чи тривалість стимуляції гормонами та іншими сигнальними молекулами функціональної активності клітин.
Система гуанілатциклаза-цГМФ-протеїнкіназа G може самостійно, без включення фосфоінозитидної системи, передавати гормональний сигнал усередину клітин. Такий механізм реалізується при зв'язуванні з мембранним рецептором натрійуретичного гормону передсердя.
Таким чином, обидва механізми дії гормонів не є різко відмінними, а можуть взаємодоповнюватись.
Гормони гіпоталамуса
В різних ділянках (нейронах) гіпоталамуса синтезуються гіпоталамічні регуляторні гормони – рилізинг-фактори (з англ. реліз – звільняти) або, за сучасною номенклатурою, ліберини і статини (рис. 6). За хімічною структурою це – низькомолекулярні пептиди. Гормони гіпоталамуса проникають у кров ворітної системи гіпофіза і з нею надходять в аденогіпофіз.
Рис. 6. Схема утворення гормонів гіпоталамуса та їх функцій
Виділення їх гіпоталамусом здійснюється під впливом нервових імпульсів, а також внаслідок змін концентрацій у крові певних гормонів (за принципом зворотного зв'язку). Ліберини стимулюють секрецію гормонів гіпофіза, а статини – гальмують. Для гіпоталамічних гормонів виявлено "перикривання ефектів", наприклад, тиреоліберин стимулює секрецію не тільки ТТГ, а і пролактину; соматостатин гальмує секрецію, крім гормону росту, також ТТГ, інсуліну, глюкагону, гастрину, секретину. Водночас соматостатин пригнічує секрецію соляної кислоти у шлунку, панкреатичного соку, перистальтику ШКТ, впливає на ЦНС. Соматостатин відкритий у різних відділах мозку, утворюється також D-клітинами острівців Лангерганса підшлункової залози, клітинами епітелію шлунка і кишечника. Механізм численних ефектів соматостатину інтенсивно вивчається.
Гіпоталамічні гормони використовуються у клінічній практиці для диференціальної діагностики гіпофізарної і гіпоталамічної недостатності (тиреоліберин, кортиколіберин), а соматоліберин – для лікування дітей із затримкою росту внаслідок недостатньої придукції соматотропіну.
Гормони гіпофіза
Функція гіпофіза така: контролює вироблення гормону щитоподібною залозою; стимулює функцію надниркових залоз; контролює функцію чоловічих і жіночих статевих залоз, а саме виділення статевих гормонів; плаценти як тимчасової залози; початок пологів, а також ріст тіла і водний баланс.
Гіпофіз може отримувати сигнали, що оповіщають про те, що відбувається в тілі, але він не має прямого зв'язку з зовнішнім середовищем. Про зовнішні впливи організм дізнається через органи чуттів, які передають отриману інформацію до центральної нервової системи. Будучи основною залозою ендокринної системи, гіпофіз сам підпорядковується центральній нервовій системі і зокрема гіпоталамусу.
Цей вищий вегетативний центр постійно координує, регулює діяльність різних відділів мозку, всіх внутрішніх органів. Частота серцевих скорочень, тонус кровоносних судин, температура тіла, кількість води в крові і тканинах, накопичення або витрату білків, жирів, вуглеводів, мінеральних солей - словом існування нашого організму, сталість його внутрішнього середовища знаходиться під контролем гіпоталамуса.
Розрізняють гормони передньої, проміжної і задньої частини гіпофіза (рис. 7).
Рис. 7. Гіпофіз: А – розташування гіпофізу; Б – схематичне зображення будови гіпофізу: 1 – передня частка (аденогіпофіз); 2 – проміжна частка; 3 – задня частка (нейрогіпофіз); 4 – епендима
Синтез гормонів передньої частки аденогіпофіза і виведення її у кров запускається ліберинами гіпоталамуса через аденілатциклазну систему. Аденогіпофіз – це не одна залоза, а комплекс залоз, кожна з яких складається з особливого типу клітин і секретує свій гормон (рис. 8). За хімічною структурою гормони аденогіпофіза відносяться до білково-пептидних: АКТГ-поліпептид; соматотропін і пролактин – прості білки, а ТТГ, ФСГ і ЛГ – складні білки (глікопротеїни). До білкової частини останніх входять 2 субодиниці, а вуглеводні ланцюги закінчуються залишками сіалової кислоти. При їх відщепленні гормони захоплюються клітинами печінки і там розпадаються
Задня частина є похідною від нервової системи (нейрогіпофіз), і в ній гормони не утворюються, а надходять по аксонах нервової клітини із гіпоталамуса. Тут вони депонуються і виділяються в кров'яне русло.
Рис. 8. Схема утворення гормонів гіпофіза та їх функцій
Обидва гормони нейрогіпофіза (вазопресин і окситоцин) за хімічною структурою є низькомолекулярними пептидами, як і гіпоталамічні ліберини і статини.
Соматотропін (соматотропний гормон (СТГ), гормон росту (ГР)
Соматотропіни є видоспецифічними білками, тому біологічна дія тваринних соматотропінів у людей не проявляється. ГР людини складається із 191 амінокислоти і містить 2 дисульфідних зв'язки. Первинна структура його визначена. Отримують соматотропін людини біотехнологічним методом. ГР виділяється гіпофізом безперервно протягом всього життя організму. Секрецію його стимулює соматоліберин, а пригнічує соматостатин.
ГР стимулює соматичний ріст органів і тканин організму, зокрема кісток, хрящів, м'язів. В основі його дії лежить вплив на обмін речовин, що здійснюється в 3-х напрямках:
1. Надходження амінокислот із крові в тканини і синтез білка, пригнічення катаболізму білків і амінокислот. Під дією ГР підвищується синтез РНК і ДНК.
2. ГР стимулює ліполіз жирів у жировій тканині, підвищує рівень жирних кислот у крові і їх утилізацію в тканинах. При тривалій дії надлишку ГР розвиваються кетоз, ожиріння печінки.
3. ГР знижує утилізацію глюкози для продукції енергії, частково завдяки підвищеній мобілізації і розпаду жирних кислот. Надходження глюкози в клітини при дії ГР короткочасно (0,5-1 год) стимулюється, і в цей період синтезується глікоген, але далі ефект ГР змінюється на протилежний і транспорт глюкози через мембрани у клітини знижується, а вміст її у крові зростає (діабетогенна дія гормону росту).
Соматотропін стимулює ріст хрящів і кісток не безпосередньо, а через стимуляцію утворення групи поліпептидів. Спочатку їх називали соматомединами, а зараз – інсуліноподібними факторами росту (ІФР). Їх концентрація у сироватці крові залежить від ГР. Найбільш вивчений ІФР‑1 (соматомедин С), який складається із 70 амінокислот. Основним місцем його синтезу вважають печінку. Біологічні ефекти ІФР-1 у хрящовій тканині такі:
1) стимуляція включення сульфату в протеоглікани;
2) стимуляція включення тимідину в ДНК;
3) стимуляція включення проліну в колаген;
4) зростання синтезу РНК і ДНК;
5) мітогенна активність, тобто стимуляція поділу клітин.
Мітогенна активність ІФР-1 проявляється і в культурах клітин інших типів, крім хрящових. Мембранні рецептори ІФР близькі за структурою до інсулінових рецепторів, володіють протеїнкіназною активністю і передають гормональний сигнал всередину клітини, стимулюють процеси транскрипції і трансляції. Взаємозв'язки гормон росту – ІФР ще вивчені недостатньо. Невідомо, які із ефектів ГР зв'язані зі стимуляцією продукції ІФР, а які – із дією самого ГР. Безпосередньо ГР впливає на транспорт амінокислот і ліполіз.
При вродженому недорозвитку гіпофіза розвивається гіпофізарна карликовість (рис. 9). Для лікування використовують ГР. У людей із мутацією, що призводить до карликовості Ларона, спостерігається високий рівень ГР у плазмі при низькому вмісті ІФР-1. У таких хворих лікування гормоном росту не стимулює ріст. Карликовість також може бути одним із проявів гіпотиреозу (кретинізму) внаслідок недостатньої секреції передньою частиною гіпофіза тиреотропного гормону. На відміну від цієї патології, гіпофізарні карлики не відстають у розумовому розвитку і не мають ознак деформації скелета.
Рис.9. Карликовість
Надмірна продукція ГР у періоді до статевого дозрівання і до завершення окостеніння зумовлює гігантизм – ріст 210‑240 см і більше, непропорційно довгі кінцівки (рис. 10).
Рис. 10. Гігантизм
У дорослих при гіперфункції гіпофіза розвивається акромегалія: непропорційно інтенсивний ріст окремих частин тіла (пальців рук і ніг, носа, нижньої щелепи, язика, внутрішніх органів) (рис. 11). Причиною акромегалії звичайно є пухлина аденогіпофіза.
Рис. 11. Обличчя хворого на акромегалію
Пролактин
За хімічною будовою – простий білок, подібний до соматотропіну. Основна функція пролактину – стимуляція утворення молока в жінок, зокрема активація синтезу білків молока (казеїну, лактальбуміну), стимуляція поглинання глюкози тканиною молочної залози і синтезу лактози, жирів.
Пролактин стимулює утворення і секрецію молока, а окситоцин – виділення молока при годуванні грудьми. Під час вагітності статеві гормони естрогени і прогестерон перешкоджають початку лактації, блокуючи дію пролактину на молочні залози. Після відторгнення плаценти при пологах і зниження рівня прогестерону зникає гальмування секреції і дії пролактину.
У плаценті виробляється подібний гормон – плацентарний лактоген людини, або соматомамотропін, який стимулює надходження глюкози в організм плода від периферичних тканин матері.
Кортикотропін (кортикотропний гормон, КТГ)
Кортиколіберин гіпоталамуса індукує транскрипцію гена проопіомеланокортину в клітинах аденогіпофіза і секрецію кортитропіну у кров. Спостерігаються добові коливання секреції, а при стресі – різке її зростання. Під контролем КТГ знаходиться пучкова зона кори надниркових залоз, клітини якої продукують кортизол. Швидкість секреції гіпофізом КТГ регулюється за принципом зворотного зв'язку рівнем кортизолу в організмі. Кортикостероїди знижують секрецію КТГ двома способами:
1) пригнічують секрецію кортиколіберину в гіпоталамусі;
2) діють безпосередньо на гіпофіз, де інгібують транскрипцію гена проопіомеланокортину.
Гальмування секреції КТГ кортизолом може перекриватись іншою регуляторною системою, більш потужною, що діє при стресі. За цих умов секреція КТГ стимулюється, незважаючи на те, що рівень кортизолу в крові високий. Механізми ще мало вивчені. Принципово важливим моментом є те, що всі нервові шляхи, які передають сигнали про біль, емоції, кровотечу, гіпоглікемію, холод, інтоксикацію хімічними речовинами і йдуть від різних ділянок головного мозку, замикаються на нейронах гіпоталамуса, які секретують кортиколіберин, і запускають стереотипну реакцію:
Рецептори КТГ розміщені на плазматичній мембрані клітини пучкової зони кори надниркових залоз. Його дія опосередковується через цАМФ і протеїнкінази. Останні активують ряд ферментів, які беруть участь у синтезі глюкокортикостероїдів. При тривалій дії КТГ на клітини надниркових залоз спостерігаються їх гіпертрофія і гіперплазія. На рівні цілого організму КТГ викликає ті реакції, які характерні для дії кортикостероїдів. Однак КТГ і безпосередньо впливає на тканини, зокрема проявляє меланоцитостимулювальну активність, ліполітичну дію в жировій тканині. При недостатньому утворенні КТГ спостерігається вторинна гіпофункція кори надниркових залоз. При пухлинах гіпофіза може мати місце гіперпродукція КТГ. Цікаво, що КТГ може синтезуватись і в пухлинних клітинах при деяких формах раку легень, аденокарциномі товстої кишки. У всіх цих випадках розвивається гіперактивність клітин кори надниркових залоз (хвороба Іценко-Кушинга).
Основні симптоми хвороби Іценко-Кушинга: ожиріння (жир відкладається в ділянці плечового пояса, над шийними хребцями и на животі); заокруглення лиця («місяцеподібне» лице); багряно-червоні щоки; тонка, суха, ціанотична шкіра; розтяжки червоно-фіолетового кольору на бедрах, грудях, плечах і животі; гематоми, що виникають при незначних травмах; гіперпігментація шкіри в області шиї, ліктів; гірсутизм у жінок (появляються вуса, борода, бакенбарди, волосся на грудях) (рис. 12); порушення менструального циклу; зниження потенції у чоловіків; гінекомастія у чоловіків (збільшення молочних залоз); підвищення артеріального тиску; атрофія мязів; збільшення живота за рахунок атрофії мязів передньої черевної стінки; акне (гнійнички на шкірі); грибкові пошкодження шкіри і нігтів; трофічні язви на ногах; болі в ділянці хребта; спонтанні переломи ребер і хребців.
Рис. 12. Гірсутизм у жінок
Меланоцитостимулювальний гормон (МСГ)
При гідролізі проопіомеланокортину утворюється також і меланоцитостимулювальний гормон. У деяких видів ссавців МСГ утворюється в клітинах проміжної частини гіпофіза. У постнатальному періоді в людини проміжна частина гіпофіза практично відсутня і МСГ у крові не визначається. На одній із стадій розвитку плід людини має виразну проміжну частину гіпофіза, в якій міститься велика кількість МСГ.
Вазопресин (антидіуретичний гормон, АДГ) і окситоцин
Ці два гормони синтезуються у тілах нейронів гіпоталамуса, по аксонах переміщаються до задньої частини гіпофіза і через нервові закінчення виділяються у кров. За хімічною природою – пептиди, утворюються із більших білків-попередників.
Дія вазопресину характеризується такими ефектами:
1. Антидіуретична дія. У клітинах ниркових канальців взаємодія АДГ з V2-рецепторами викликає підвищення рівня цАМФ, фосфорилювання поки що невідомих білків, що зумовлює збільшеня проникності мембрани для води, і реабсорбцію води, вільної від іонів, за градієнтом концентрації із гіпотонічної первинної сечі через клітини в позаклітинну рідину. В результаті осмотичний тиск плазми крові і тканинної рідини зменшується і секреція гормону припиняється.
2. Підтримка артеріального тиску. Взаємодія АДГ з V1-рецепторами гладком'язових клітин в судинах викликає збільшення концентрації іонів кальцію в клітинах і скорочення м'язів, звуження судин, підвищення кров'яного тиску. Пресорний ефект вазопресину спостерігається при дії значної кількості гормону.
3. Участь у механізмах пам'яті. АДГ позитивно діє на закріплення пам'яті й мобілізацію інформації, що зберігається. Клітинні механізми впливу АДГ на ЦНС вивчені недостатньо.
Секреція АДГ регуюється змінами осмотичного тиску і об'єму циркулюючої крові, а також різними нейрогенними стимулами. Специфічні осморецептори мозку реагують на підвищення осмотичного тиску плазми крові і тканинної рідини сигналами про виділення вазопресину в кров і навпаки. При крововтратах, зниженні об'єму крові барорецептори клітин кровоносних судин передають сигналами в ЦНС і стимулюють секрецію АДГ, а також альдостерону. Вивільнення АДГ гальмується адреналіном.
При недостатності АДГ виникає нецукровий
діабет, при якому за добу із організму виводиться 10-
Окситоцин проявляє 2 біологічні ефекти: скорочення мускулатури матки і виділення молока. Концентрація рецепторів до окситоцину в гладкій мускулатурі матки зростає під час вагітності і досягає максимуму на ранній стадії родового акту. Естрогени сенсибілізують міометрій до дії окситоцину, а прогестерон знижує. Окситоцин бере участь у початку родів як безпосередньо, викликаючи скорочення м'язів матки, так і опосередковано, стимулюючи утворення простагландинів, які є сильним активаторами скорочення гладких м'язів.Окситоцин використовується у клініці для стимуляції родів. Виділення молока окситоцином стимулюється внаслідок скорочення м'язових волокон, розміщених навколо альвеол молочних залоз.
Гормони підшлункової залози
Ендокринні клітини острівців Лангерганса підшлункової залози синтезують ряд гормонів: А-клітини – глюкагон, В-клітини – інсулін, D‑клітини – соматостатин, F-клітини – панкреатичний поліпептид. Біологічна роль останнього мало вивчена (рис. 13).
Рис. 13. Схема будови ендокринної частини підшлункової залози
Недостатність інсуліну зумовлює розвиток цукрового діабету. Історія вивчення діабету, підшлункової залози й інсуліну відображає, по суті, всю історію біології і медицини.
Інсулін
Швидкість секреції інсуліну залежить від концентрації глюкози в крові. При нормальному рівні глюкози в крові натще (3,33-5,5 ммоль/л) секреція інсуліну мінімальна. Під час споживання їжі підвищення концентрації глюкози в крові викликає збільшення секреції інсуліну. Механізм регуляторного впливу глюкози на секрецію інсуліну досить складний і зв'язаний зі швидкістю транспорту іонів Са2+ через плазматичну мембрану В-клітин і інтенсивністю гліколізу в них. Конкретний метаболіт глюкози, який активує секрецію інсуліну, поки що невідомий. На швидкість синтезу і секреції інсуліну впливають також гормон росту, глюкагон, адреналін, секретин, холецистокінін, соматостатин, причому, за винятком адреналіну і соматостатину, всі інші збільшують секрецію інсуліну.
Біологічні ефекти інсуліну. Рецептори інсуліну відкриті в багатьох типах клітин. Головними мішенями дії інсуліну є клітини м'язів, печінки, жирової тканини. Рецептори локалізовані у плазматичній мембрані, за хімічною природою є глікопротеїнами, вуглеводна частина яких знаходиться на зовнішній стороні мембрани. Рецептор складається із 4 субодиниць: дві альфа‑субодиниці зв'язують інсулін, а дві бета-субодиниці є трансмембранними білками з активністю тирозинкінази. При зв'язуванні інсуліну з рецептором стимулюється кіназна активність бета-субодиниць і відбувається автофосфорилювання їх, а також фосфорилювання ряду інших білків, що, у свою чергу, індукує активність цілого ряду ферментів. Вірогідно існує декілька вторинних посередників дії інсуліну, зокрема продукти розпаду інозитфосфатидів. Таким чином, інсулін запускає багатокаскадну розгалужену систему регуляторних реакцій.
Дія інсуліну на обмін вуглеводів. 1. Підвищення перенесення глюкози з крові в клітини м'язів, жирової тканини, лімфатичної тканини, печінки тощо. Під впливом інсуліну надходження глюкози в клітини м'язів, що знаходяться в стані спокою, зростає у 15-20 разів. Надходження глюкози в мозок, нерви, мозковий шар нирок, зародковий епітелій сім'яників, клітини ендотелію судин, кришталик не залежить від інсуліну. Точний механізм активації інсуліном транспортної системи для глюкози невідомий.
2. Активація глюкокінази, глікогенсинтетази печінки і в результаті збільшення синтезу глікогену. Також зростає синтез глікогену в м'язах. Інсулін гальмує дію адреналіну і глюкагону на процес глікогенолізу, знижуючи вміст у клітинах цАМФ.
3. Стимуляція гліколізу і використання продуктів розпаду (діоксіацетонфосфату і ацетил-КоА) для синтезу жирів. При тривалій дії інсулін індукує синтез ключових ферментів глюколізу.
4. Гальмування глюконеогенезу завдяки зниженню активності регуляторних ферментів процесу і пригнічення надходження амінокислот із позапечінкових тканин у печінку.
Отже, інсулін пригнічує утилізацію жирів і стимулює їх синтез. Можна зробити висновок, що одна із важливих функцій інсуліну полягає у зміні катаболізму вуглеводів і жирів для забезпечення організму енергією. При високій концентрації глюкози інсулін включає утилізацію вуглеводів і гальмує катаболізм жирів. І навпаки, при низькій концентрації глюкози низький вміст інсуліну в крові викликає утилізацію жиру в усіх тканинах, крім мозку.
Дія інсуліну на обмін білків і нуклеїнових кислот.
1. Стимуляція транспорту амінокислот із крові в тканини.
2. Підвищення синтезу білків у тканинах завдяки збільшенню концентрації амінокислот і стимуляції процесу трансляції матричних РНК.
3. Гальмування катаболізму білків, виходу амінокислот із тканини у кров.
4. Інсулін стимулює синтез ДНК і РНК. Збільшення швидкості реплікації і транскрипції забезпечує проліферацію клітин.
Таким чином, інсулін стимулює синтез білків і нуклеїнових кислот, зумовлює позитивний азотний баланс. Разом із соматотропіном інсулін стимулює ріст організму.
Цукровий діабет
Розрізняють інсулінозалежні та інсулінонезалежні форми цукрового діабету. У першому випадку рівень інсуліну в крові значно нижчий, ніж у нормі, а у другому випадку рівень інсуліну може знаходитись у межах норми або навіть вище. При інсулінонезалежному діабеті має місце інсулінорезистентність клітин-мішеней, тобто зниження відповіді їх на ендогенний і екзогенний інсулін. У деяких випадках резистентність до інсуліну є наслідком зменшення кількості рецепторів до інсуліну.
Характерні для цукрового діабету біохімічні зміни і клінічні симптоми є наслідком основної причини – абсолютної чи відносної недостачі інсуліну, яка супроводжується відносним надлишком глюкагону і глюкокортикоїдів.
Вуглеводний обмін при цукровому діабеті характеризується такими ознаками:
1) зниження надходження глюкози з крові в тканини;
2) гальмування синтезу і стимуляція розпаду глікогену в печінці;
3) зростання глюконеогенезу з амінокислот і гліцерину.
У результаті концентрація глюкози в крові зростає (гіперглікемія). Рівень гіперглікемії натщесерце у комплексі з іншими ознаками використовується для визначення ступеня тяжкості цукрового діабету. При прихованому діабеті концентрація глюкози в крові натщесерце перебуває у межах норми. Для виявлення прихованих порушень обміну вуглеводів використовують метод цукрового навантаження (тест на толерантність до глюкози). Через 2 години після навантаження при явному і при прихованому діабеті концентрація глюкози в крові чітко є вищою від вихідної (знижена толерантність до глюкози). У хворих на цукровий діабет із тяжкою формою перебігу гіперглікемія може досягати високих величин (25-30 ммоль/л і вище). При гіперглікемії вище ниркового порогу (приблизно 10 ммоль/л) глюкоза виділяється із сечею (глюкозурія).
При діабеті ліпіди забезпечують енергією тканини організму, за винятком мозку, зростає мобілізація жиру з депо, жирні кислоти транспортуються до печінки і меншою мірою до інших тканин. Окиснення їх частково забезпечує клітини енергією. Надлишок жирних кислот у печінці використовується в таких напрямках:
1) синтез кетонових тіл;
2) синтез жирів і фосфоліпідів;
3) синтез холестерину.
Синтезовані у печінці жири, фосфоліпіди і холестерин виводяться в кров у вигляді ліпопротеїнів дуже низької густини. Підвищується рівень ліпідів у крові (гіперліпопротеїнемія). При важких формах діабету швидко розвивається атеросклероз. Підвищений синтез жирів у печінці хворих на цукровий діабет також може призвести до ожиріння печінки.
Посиленому синтезу кетонових тіл сприяє надлишок у гепатоцитах ацетил-КоА, який утворюється при бета-окисненні жирних кислот (рис. 14).
Рис. 14. Збільшення кетогенезу та синтезу холестерину при підвищеному розпаді жирних кислот та гальмуванн ЦТК
Використання ацетил-КоА в циклі Кребса (цитратсинтазна реакція) сповільнюється через дефіцит оксалоацетату, який за цих умов використовується для глюконеогенезу. Синтез жирних кислот із ацетил-КоА блокується внаслідок гальмування надлишком жирних кислот початкової реакції процесу – утворення малоніл-КоА (ацетил-КоА-карбоксилазної реакції). Дефіцит НАДФН також зумовлює гальмування синтезу жирних кислот. Таким чином, залишається незаблокованою тільки конденсація ацетил-КоА до бета-гідрокси-бета-метилглутарил-КоА з подальшим утворенням кетонових тіл і холестерину.
Кетонові тіла, ацетооцтова і бета-оксимасляна кислоти переносяться кров'ю до позапечінкових тканин, де окиснюються з виділенням енергії (рис. 15).
Рис. 15. Схема синтезу і розпаду кетонових тіл
Але швидкість утилізації кетонових тіл у тканинах відстає від швидкості їх утворення через дефіцит оксалоцетату.
Значно зростає концентрація кетонових тіл у крові (кетонемія), виведення їх із сечею (кетонурія). Разом з ацетооцтовою і бета-оксимасляною кислотами виводяться з організму іони натрію, що використовуються для нейтралізації цих кислот.
З ацетооцтової кислоти внаслідок спонтанного декарбоксилювання утворюється ацетон, який виділяється легенями. Три симптоми – кетонемію, кетонурію і запах ацетону при диханні – об'єднують під спільною назвою “кетоз”. Ацетооцтова і бета-оксимасляна кислоти відносяться до помірно сильних кислот, і зростання їх вмісту в крові зумовлює розвиток метаболічного ацидозу. рН крові падає до 7,1-7,0 і навіть нижче.
Відсутність інсуліну викликає зниження синтезу і зростання розпаду білків у тканинах. Амінокислоти частково окиснюються і служать джерелом енергії, а частково надходять у печінку і використовуються для глюконеогенезу. Внаслідок підвищення утилізації амінокислот зростає утворення сечовини і виведення її з організму. Підвищується рівень залишкового азоту в крові (азотемія) й азоту в сечі (азотурія). Розвиваються атрофія м'язів, кахексія.
Виведення через нирки значної кількості глюкози, кетонових тіл, сечовини, іонів Nа+ супроводжується втратою рідини, оскільки осмотичний тиск цих речовин у первинній сечі перешкоджає реабсорбції води в ниркових канальцях (осмотичний діурез). Добовий об'єм сечі може зростати у декілька раз (поліурія). Підвищується виведення з сечею Na+, K+ та інших електролітів. Розвивається зневоднення (дегідратація) організму і як наслідок його – посилена спрага (полідипсія). Різке падіння об'єму циркулюючої крові призводить до недостатності периферичного кровообігу і гіпоксії тканин. У результаті ацидозу, гіпоксії і клітинної дегідратації порушується функція мозку з втратою свідомості (діабетична кома).
Кетоацидоз і дегідратація організму спостерігаються при вираженій гострій недостачі інсуліну. При хронічній недостачі інсуліну розвиваються ускладнення цукрового діабету – ураження нирок, стінок судин, нейропатії, катаракта.
Для лікування використовуються різні форми інсуліну. Розробляються такі методи лікування, як пересадка острівців підшлункової залози, тільки бета-клітин, використання штучної залози, яка включає аналізатор концентрації глюкози і програмовий дозатор інсуліну.
Глюкагон
Глюкагон – це поліпептид, який складається із 29 амінокислотних залишків. Синтезується з білка-попередника в А-клітинах підшлункової залози: препроглюкагон – проглюкагон – глюкагон.
Після синтезу глюкагон депонується в гранулах і вивільняється в кров шляхом екзоцитозу. Секреція глюкагону гальмується глюкозою, іонами Са2+ та інсуліном. Концентрація глюкагону й інсуліну в крові змінюється протилежним чином: відношення інсулін/глюкагон максимальне під час травлення і мінімальне при голодуванні. Додатковим фактором є характер їжі. При споживанні великої кількості білків амінокислоти стимулюють секрецію і глюкагону, і інсуліну. Секреція одного інсуліну може викликати гіпоглікемію, а одночасне звільнення глюкагону компенсує гіпоглікемічний ефект інсуліну, стимулюючи глікогеноліз і глюконеогенез. При споживанні змішаної їжі глюкоза гальмує секрецію глюкагону і попереджує його викид під дією амінокислот.
Органи-мішені для глюкагону: печінка, міокард, жирова тканина, але не скелетні м'язи. Глюкагон взаємодіє з рецепторами, які локалізовані на плазматичній мембрані, що викликає активацію аденілатциклази, збільшення рівня цАМФ і активацію протеїнкіназ. Фосфорилювання регуляторних ферментів під дією протеїнкіназ стимулює одні метаболічні процеси і гальмує інші.
Механізм дії цього гормону - мембранно-цитозольний, через активацію аденілатциклази і утворення цАМФ.
Біохімічні ефекти глюкагону протилежні ефектам інсуліну (це контрінсулярний гормон). Цей гормон викликає потужну мобілізацію джерел енергії за рахунок активації катаболізму вуглеводів і ліпідів.
Вплив на обмін вуглеводів спрямований на підвищення концентрації глюкози в крові. Основна “мішень” дії глюкагону - печінка, де він:
1) стимулює розщеплення глікогену (активує глікогенфосфо-рилазу) і інгібує глікогенез (гальмує активність глікоген-синтази); глюкагон не впливає на глікогеноліз у м’язах;
2) підвищує інтенсивність глюконеогенезу. Це досягається через активацію синтезу ключового ферменту фосфоенолпіруват-карбоксикінази (ФЕПКК) - цАМФ підвищує швидкість транскрипції гену ФЕПКК
Вплив глюкагону на обмін ліпідів полягає в активації ліполізу. В адипоцитах гормон активує гормончутливу ліпазу, що сприяє підвищенню вільних жирних кислот в крові. Ці жирні кислоти можуть бути використані як джерела енергії або перетворюватися в печінці в кетонові тіла. В умовах інсулінової недостатності цей механізм є суттєвим у розвитку кетонемії.
Таким чином, глюкагон та інсулін є функціональними антагоністами. Ефекти глюкагону – це перша лінія захисту організму від гіпоглікемії в період голодування чи підвищених енергетичних затрат. Глюкоза в цих умовах використовується мозком, а в м'язах і в інших інсулінозалежних тканинах джерелом енергії служать жирні кислоти і кетонові тіла.
Гормони мозкового шару надниркових залоз
У хромафінній тканині мозкового шару наднирників утворюються катехоламіни (адреналін - 85%, норадреналін – 15%).
Адреналін (епінефрин) - гормон, який секретується в наднирниках при стресі (концентрація його в крові зростає в 4-5 разів) і є медіатором в деяких синапсах. Встановлено також, що при стресі вміст в крові норадреналіну, який має переважно медіаторні властивості також зростає.
Біохімічні ефекти адреналіну:
Адреналін - це контрінсулярний гормон, дія якого спрямована насамперед на збільшення концентрації глюкози в крові. Крім того, цей гормон має катаболічну дію на обмін ліпідів.
Вплив адреналіну на обмін вуглеводів полягає в стимуляції глікогенолізу в печінці і м’язах. Це досягається шляхом активації глікогенфосфорилази. У м’язах відбувається розщеплення глікогену до глюкозо-6-фосфату і далі до лактату. У м’язах розщеплення глікогену до глюкози не відбувається, тому що відсутня глюкозо-6-фосфатаза (яка розщеплює глюкозо-6-фосфат до глюкози). У печінці в результаті глікогенолізу утворюється вільна глюкоза, яка надходить у кров, що призводить до підвищення рівня глікемії. Це має велике значення для забезпечення інших тканин паливом в умовах стресу.
Вплив адреналіну на обмін ліпідів полягає в стимуляції ліполізу в жировій тканині через активацію ключового ферменту цього процесу - ТАГ-ліпази. У результаті в крові підвищується вміст вільних жирних кислот, що є додатковим джерелом метаболічної енергії.
Таким чином, сумарний біохімічний ефект адреналіну - це підготовка до ситуацій, в яких організм використовує значну кількість метаболічної енергії.
В інактивації катехоламінів беруть участь два ферменти:
1) моноамінооксидаза (МАО) - мітохондріальний фермент;
2) катехол-О-метилтрансфераза - фермент цитозолю.
Найбільша активність цих ферментів спостерігається в печінці і нирках. Визначення концентрації катехоламінів в крові є важливим діагностичним підтвердженням діагнозу феохромоцитом.
Феохромоцитоми -
пухлини, які розвиваються у мозковій речовині наднирника, яка володіє здатністю
виробляти адреналін і норадреналін, що призводить до розвитку різких підйомів
артеріального тиску.
Феохромоцитома
по праву вважається самою підступною й непередбачуваною пухлиною. Розвиваються,
як правило, раптово гіпертонічні кризи, які супроводжуються серцебиттям,
панікою, тремором рук і ніг, ознобом, підвищеним потовиділенням,
запамороченням. Протягом
2-3 хвилин артеріальний тиск може досягти 250 -
Синтез катехоламінів регулюється за принципом негативного зворотного зв'язку. Норадреналін гальмує активність тирозингідроксилази, адреналін-метилтрансферази. У хромафінних клітинах мозкової речовини надниркових залоз адреналін і норадреналін накопичуються в секреторних гранулах. Їх синтез і вивільнення у кров шляхом екзоцитозу регулюються нервовими центрами, розміщеними в гіпоталамусі. Збудження симпатичної нервової системи стимулює секрецію катехоламінів, причому спочатку більше вивільняється адреналіну, а при тривалій стимуляції – норадреналіну. Надниркові залози людини містять в нормі адреналіну в 3-10 разів більше, ніж норадреналіну, а концентрація в крові норадреналіну – 5,2 нмоль/л проти 1,9 нмоль/л адреналіну, що зумовлено частковим надходженням норадреналіну в кров із синапсів при стимуляції симпатичних нервів. Вміст катехоламінів у крові зростає дуже швидко (майже у 1000 разів під час стресових реакцій). Із сечею за добу виділяється 11-76 нмоль адреналіну і 47-236 нмоль норадреналіну.
Гормони щитовидної залози
У щитовидній залозі синтезуються гормони двох груп:
1) тиреоїдні гормони, інакше йодтироніни, з дуже широким спектром дії;
2) кальцитонін, який регулює обмін кальцію і фосфатів.
Щитовидна залоза секретує у кров два тиреоїдні гормони – тироксин (тетрайодтиронін) і трийодтиронін. Скорочено вони позначаються Т4 і Т3. За хімічною природою ці гормони є похідними амінокислоти тирозину.
Для синтезу йодтиронінів необхідний мікроелемент йод у формі йодиду. Щитовидна залоза концентрує йодид із плазми крові за допомогою йодидної помпи, тобто системи активного транспорту.
Повністю йодований тиреоглобулін, який називають йодтиреоглобуліном, містить у поліпептидному ланцюзі монойодтирозин, дийодтирозин, трийодтиронін і тетрайодтиронін. Синтез відбувається у фолікулярних клітинах залози, із яких йодтиреоглобулін надходтиь у фолікули, заповнені колоїдом. Тиреоглобулін служить запасною формою тиреоїдних гормонів, Т3 і Т4. Коли на клітини щитовидної залози діє тиреотропний гормон гіпофіза, невеликі краплинки колоїду надходять із фолікула назад у клітини і лізосомні протеази гідролізують йодтиреоглобулін до амінокислот і вуглеводів. Звільнені Т3 і Т4 секретуються у кров, а монодийодтирозини можуть знову використовуватись. Для цього спеціальний фермент дегалогеназа каталізує дейодування і утворені йодид і тирозин йдуть на синтез йодтиреоглобуліну. Т3 і Т4 не піддаються дії дегалогенази.
Регуляція синтезу і секреції тиреоїдних гормонів
Регуляція здійснюється через гіпоталамо-гіпофізарну систему.
Із гіпоталамуса постійно вивільняється тиреоліберин, який через цАМФ викликає секрецію із клітин аденогіпофіза тиреотропіну (ТТГ). Гальмує цей процес соматостатин. Секреція тиреоліберину і ТТГ досить постійна і підвищується при зниженні температури навколишнього середовища. ТТГ взаємодіє з рецепторами мембрани епітеліальних клітин фолікулів та через цАМФ і, вірогідно, інші вторинні посередники стимулює синтез і секрецію тиреоїдних гормонів. При цьому відбувається захоплення йодиду щитовидною залозою, синтез тиреоглобуліну, гідроліз молекул тиреоглобуліну, що знаходились у фолікулах, секреція Т4 і Т3 у кров.
Тиреоїдині гормони необхідні для нормального росту та розвитку дитячого організму, особливо для розвитку нервової системи. Вони також впливають на метаболізм дорослого організму.
Біологічна дія тиреоїдних гормонів
Йодтироніни діють практично на всі органи і тканини організму. Розрізняють їх вплив, з одного боку, на процеси розвитку організму, диференціювання клітин, а з іншого – на основний обмін, теплопродукцію. Зокрема, тиреоїдні гормони стимулюють ріст і розвиток мозку в ембріона і протягом перших декількох років після народження. Значна недостатність гормонів у дитячому віці зумовлює затримку росту, розумову відсталість.
Білки-рецептори до тиреоїдних гормонів локалізовані і на плазматичній мембрані, і в ядрі, і в мітохондріях, і в цитоплазмі. Зв'язування гормонів із рецепторами плазматичної мембрани відіграє певну роль у транспорті їх у клітину, а також стимулює транспорт у клітини амінокислот. Рецептори цитоплазми функціонують, напевно, як внутрішньоклітинні переносники гормонів. Ядерні білки-рецептори проявляють значно більшу спорідненість із Т3. Комплекс гормону з ядерними рецепторами викликає активацію процесу транскрипції, збільшення синтезу певного набору матричних РНК, а також рибосомної РНК. У результаті підвищується синтез таких ферментів, як Na+, К+-АТФаза, мітохондріальні ферменти тканинного дихання, глюкозо-6-фосфатдегідрогеназа, НАДФ-малатдегідрогеназа, гліцерофосфатдегідрогеназа, ферментів ліпогенезу і ліполізу. Синтез білків забезпечується амінокислотами, надходження у клітини яких зростає під дією тиреоїдних гормонів.
Головний результат дії тиреоїдних гормонів полягає у зростанні швидкості основного обміну, окисненні вуглеводів, жирів, амінокислот. Підвищується споживання кисню і виділення СО2. Механізм процесів, що лежать в основі підвищення тиреоїдними гормонами теплопродукції (калоригенного ефекту), досліджується протягом довгого часу. Раніше вважали, що калоригенний ефект зумовлюється роз'єднанням під дією Т3 процесів тканинного дихання і окиснювального фосфорилювання, тому що гормон підвищує споживання кисню і окиснення субстратів без відповідного збільшення синтезу АТФ. Але ця точка зору не отримала підтвердження в дослідженнях. Більш вірогідним поясненням підвищення теплопродукції під дією тиреоїдних гормонів вважають збільшення використання АТФ в енергозалежних процесах, зокрема на активне перенесення іонів Na+ і К+. Використання АТФ зумовлює збільшення вмісту АДФ, що стимулює процеси катаболізму білків, жирів, вуглеводів і синтезу АТФ. Підвищений вміст тиреоїдних гормонів викликає одночасну стимуляцію протилежно спрямованих процесів, наприклад ліпогенезу (за рахунок індукції синтезу ферментів ліпогенезу) і ліполізу, синтезу білків і розпаду їх з окисненням амінокислот. У результаті енергія, використана на процеси синтезу, розсіюється внаслідок прискорення катаболізму, що зумовлює підвищення теплопродукції. Необхідно сказати, що функції щитовидної залози давно пов'язують з адаптацією до низьких температур.
Тиреоїдні гормони стимулюють захоплення клітинами глюкози, гліколіз і глюконеогенез, мобілізацію жиру із жирового депо, окиснення жирних кислот, синтез холестерину і перетворення його в жовчні кислоти. Під впливом Т4 в крові знижується концентрація холестерину, ліпопротеїнів, але підвищується вміст вільних жирних кислот. Підвищена концентрація тироксину активує глюкозо-6-фосфатазу, що призводить до розвитку гіперглікемії. Тиреоїдні гормони підвищують кровообіг, особливо у шкірі для відведення тепла, частоту скорочень серця, глибину дихання.
Таким чином, дія гормонів щитовидної залози різноспрямована і не однозначна. Крім того, дія Т3 і Т4 залежить від їх концентрації в крові. У фізіологічних концентраціях вони стимулюють анаболічні процеси при позитивному азотовому балансі. При підвищених концентраціях тиреоїдних гормонів переважають катаболічні процеси.
Тиреоїдні гормони інактивуються шляхом дейодування, дезамінування, деградації бокового ланцюга. Самі гормони і деякі їх метаболіти утворюють кон’югати з глюкуроновою кислотою і, рідше, з сірчаною. Більшість реакцій метаболізму тиреоїдних гормонів відбувається у печінці. Кінцеві продукти виділяються з жовчю. Йодид знову надходить у щитовидну залозу, а частина його екскретується з сечею. Гіпотиреоз у дитячому віці зумовлює затримку росту і непропорційний ріст тіла, затримку психічного розвитку, кретинізм.
Гіпотиреоз у дорослих характеризується такими змінами:
1) зниження активності ферментів, споживання тканинами кисню і, таким чином, зменшення основного обміну, теплопродукції і підвищення чутливості до холоду;
2) порушення водно-сольового обміну – розвивається слизовий набряк (мікседема) внаслідок сповільнення обміну протеогліканів і глікопротеїнів сполучної тканини, що сприяє затримці води, особливо у підшкірній клітковині;
3) порушення з боку серцево-судинної системи, м'язів, ШКТ, послаблення імунітету;
4) гіперхолестеринемія (гіпербеталіпопротеїнемія), що сприяє розвитку атеросклерозу;
5) порушення на рівні нервової системи – психічна і фізична в'ялість, сонливість.
Більшість видів гіпотиреозу лікується тиреоїдними гормонами.
Однією із форм гіпофункції щитовидної залози є ендемічний зоб, що розвивається при недостатньому надходженні в організм йоду в певних районах, де низький його вміст у воді і грунтах. При ендемічному зобі низька концентрація в крові тиреоїдних гормонів призводить за принципом зворотного зв'язку до підвищеної секреції тиреотропіну гіпофіза, який стимулює розростання щитовидної залози до таких розмірів, поки не встановиться рівновага між розмірами залози і використанням невеликої кількості йоду. Такий компенсаторний механізм до певного часу забезпечує достатню концентрацію тиреоїдних гормонів і рівень основного обміну. Але далі переважає розростання сполучної тканини залоз і розвивається гіпотиреоз. Для попередження розвитку ендемічного зоба кухонну сіль збагачують йодидами, вживають морські продукти.
Гіперфункція щитоподібної залози
При гіпертиреозі (дифузному токсичному зобі, або Базедовій хворобі чи хворобі Грейвса) мають місце підвищений синтез і секреція Т3 і Т4, спостерігаються збільшення розмірів залози (зоб), екзофтальм, підвищення основного обміну на 30-60 % вище норми, тахікардія, підвищення сили скорочень серця (при вираженій гіперфункції сила скорочень знижується внаслідок посиленого катаболізму білків міокарда), м'язова слабкість, підвищені апетит і споживання їжі, але одночасно втрата маси тіла внаслідок переважання катаболізму, розвиток психоневротичних порушень.
Причинами дифузного токсичного зоба є автоімунні процеси.
Дифузний токсичний зоб
Може розмножуватися клон клітин, які синтезують антитіла до білків-рецепторів тиреотропного гормону в клітинах щитоподібної залози. Дія таких антитіл аналогічно до дії ТТГ стимулює синтез і секрецію тиреоїдних гормонів. Іншою причиною гіпертиреозу може бути аденома гіпофіза і, в результаті, підвищена продукція ТТГ.
Гормональний контроль обміну кальцію
Концентрація Са2+ в плазмі крові регулюється з дуже великою точністю, зміна її всього на декілька відсотків зумовлює дію гомеостатичних механізмів для відновлення фізіологічного рівня (приблизно 2,5 ммоль/л). У підтримці гомеостазу Са2+ беруть участь три основних гормони: паратгормон, кальцитонін і активна форма вітаміну D3 – 1,25-дигідроксихолекальциферол, який функціонує як гормон.
Утворення і секреція паратгормону та кальцитоніну в кров регулюється концентрацією іонного кальцію в плазмі. При нормальній концентрації Са2+ секретуються невеликі кількості обох гормонів. При зниженні концентрації Са2+ паращитоподібні залози збільшують секрецію паратгормону, а секреція кальцитоніну гальмується. При підвищенні концентрації Са2+ виділяється кальцитонін, а активність паращитоподібних залоз знижується.
Під впливом паратгормону концентрація Са2+ в плазмі крові зростає, що зумовлюється його дією на кістки, нирки і кишечник. Плазматичні мембрани клітин цих органів містять рецептори, які зв'язують паратгормон. Ця взаємодія активує аденілатциклазну систему. В кістках стимулюється остеоліз (резорбція) остеокластами і остеоцитами, вивільнення іонів кальцію і фосфатів у кров. Разом із цим розпадається органічний матрикс, на що вказує зростання екскреції із сечею гідроксипроліну – маркера колагену. При дії паратгормону також гальмується синтез колагену в активних остеобластах. Тривалий вплив паратгормону викликає збільшення кількості остеокластів, а потім і остеобластів. Дійсний механізм вивільнення Са2+ із мінеральної структури кісток залишається невідомим. Припускають, що у відповідь на дію паратгормону в кістках накопичуються органічні кислоти (лактат, цитрат, ізоцитрат), що створює оптимум рН для дії протеолітичних ферментів, можливо, лізосомного походження.
Значну роль у підвищенні концентрації Са2+ в плазмі крові відіграє дія паратгормону на нирки, де він стимулює реабсорбцію Са2+ дистальними канальцями. Одночасно паратгормон гальмує реабсорбцію фосфатів, що призводить до зниження їх вмісту у крові і фосфатурії.
Другим важливим ефектом паратгормону є стимуляція синтезу 1,25-дигідроксихолекальциферолу, активної форми вітаміну D, який стимулює всмоктування кальцію в тонкій кишці. Механізм дії 1,25-дигідроксихолекальциферолу, який має стероїдну структуру, такий же, як у стероїдних гормонів. У клітинах тонкої кишки він індукує синтез ряду білків, серед яких є кальцієзв'язувальний білок, здатний переносити іони Са2+. 1,25(ОН)2D3 також підвищує всмоктування в тонкій кишці фосфатів. Таким чином, дія вітаміну D спрямована на забезпечення безперервного надходження в організм мінеральних речовин для відкладання в кістковій тканині. Метаболіти вітаміну D діють і безпосередньо на клітини кісток, стимулюючи синергічно з паратгормоном мобілізацію мінеральних компонентів із кісткової тканини.
Кальцитонін гальмує резорбцію кісткової тканини остеокластами і остеоцитами, що супроводжується зниженням вмісту в плазмі крові кальцію і фосфатів. Дія кальцитоніну реалізується через аденілатциклазну систему. При тривалій дії кальцитоніну зменшується утворення остеокластів із клітин-попередників, що вторинно викликає зменшення числа остеобластів. У нирках збільшується екскреція фосфатів, напевно, внаслідок зміни концентрації Са2+ в плазмі. Таким чином, на виведення фосфатів кальцитонін і паратгормон діють як синергісти, а на концентрацію кальцію в крові і на кістки – як антагоністи.
Крім паратгормону, активної форми вітаміну D і кальцитоніну, на метаболізм кісткової тканини і вміст кальцію та фосфатів у рідинах впливають інші гормони. Зокрема, виражену дію на ріст кісток проявляють гормони росту, тиреоїдні гормони, естрогени і андрогени, простагландини.
Гормони кіркової речовини надниркових залоз
Кортикостероїди відносяться до групи стероїдних гормонів, які синтезуються з холестерину.
Загальна схема синтезу стероїдних гормонів: надниркові залози:
пучкова зона – К, Кк; клубочкова зона – Ал; сітчаста зона – Ан; сім’яники – Т; яєчник – Е; жовте тіло – П, Е; плацента – П, Е.
Гормони ліпофільної природи (стероїдні гормони), молекули яких здатні проникати через ліпідній бішар плазматичної мембрани клітин, проявляють свою дію за допомогою цитозольного механізму. До них відносять всі стероїдні гормони, а також йодтироніни, які за ліпофільністю займають проміжне положення між стероїдами та водорозчинними гормонами. Специфічні рецептори для цих гормонів містяться в цитоплазмі тільки клітин-мішеней. Вони є білками, що мають високу спорідненість до свого гормону за рахунок стереоспецифічності. Коли стероїдний або тиреоїдний гормон проникає через плазматичну мембрану в середину клітини, в цитоплазмі він зв'язується з цитозольним рецептором, утворює комплекс гормон – рецептор, який надалі зазнає активації. У процесі активації змінюється конформація, розміри та поверхневий заряд комплексу, він виявляє здатність проникати в ядро і зв'язуватися з певними ділянками хроматину, активувати або інактивувати специфічні гени. Внаслідок впливу на транскрипцію генів змінюється вміст певних білків, що позначається на активності біохімічних процесів у клітині. Для стероїдних та тиреоїдних гормонів, які здійснюють прямий вплив на генетичний апарат клітини, більш характерна регуляція росту та диференціювання тканин і органів, ніж для гормонів, що не проникають у клітину.
Залежно від біологічної дії кортикостероїди поділяють на глюкокортикоїди, що впливають на обмін органічних речовин, і мінеролокортикоїди, що впливають на водно-сольовий обмін. Глюкокортикоїди – кортизол і кортизон. Мінералокортикоїди – альдостерон і дезоксикортикостерон. Кортикостерон проявляє властивість обох груп, але значно менш активний. Відзначимо, що глюкокортикоїди синтезуються головним чином у пучковій зоні кори надниркових залоз, альдостерон – у клубочковій, а клітини сітчастої зони виробляють кортикостероїди з андрогенною активністю.
Біологічна дія глюкокортикоїдів
Мішенями для дії глюкокортикоїдів є більшість органів і тканин, зокрема печінка, м'язи, нирки, кістки, шкіра, жирова і лімфоїдна тканини. Прямо або опосередковано глюкокортикоїди регулюють більшість фізіологічних і біохімічних процесів. В одних тканинах вони стимулюють процеси катаболізму, в інших – процеси анаболізму. Вважають, що і катаболічні, і анаболічні реакції зумовлюються впливом глюкокортикоїдів на транскрипцію генів. Як і всі стероїдні гормони, кортикостероїди легко проходять через плазматичну мембрану клітин-мішеней у цитоплазму, де з'єднуються із специфічними білками-рецепторами. Розрізняють рецептори глюкокортикоїдів типу I i II. Гормон-рецепторний комплекс проникає у ядро, де зв'язується з певними ділянками ДНК і індукує транскрипцію певних генів.
В результаті стимуляції глюконеогенезу глюкокортикоїди збільшують концентрацію глюкози в крові і синтез глікогену в печінці. Крім того, глюкокортикоїди зменшують використання глюкози у периферичних тканинах (скелетних м'язах, жировій і лімфоїдній тканинах). При високому рівні глюкокортикоїдів значне підвищення глюконеогенезу і зниження утилізації глюкози клітинами призводять до розвитку стану, який називається стероїдним (адреналовим) діабетом: гіперглікемії, глюкозурії, кетонемії, діабетичного типу кривої при цукровому навантаженні.
Зовнішній вигляд хворої, що страждає на синдром Іценко – Кушінга
Із гальмуванням утилізації глюкози зв'язана ліпідомобілізуюча дія глюкокортикоїдів. Вони підвищують реакцію жирових клітин на ліполітичні гормони, збільшують вміст вільних жирних кислот у плазмі і покращують утилізацію їх клітинами, стимулюють утворення кетоновий тіл. У печінці, на відміну від жирової тканини, глюкокортикоїди збільшують синтез жирів.
Глюкокортикоїди і їх синтетичні аналоги з успіхом використовуються у клінічній практиці як протизапальні препарати, а також при лікуванні важких алергічних станів і, разом з імуносупресорами, для профілактики відторгнення трансплантатів.
Біологічна дія мінералокортикоїдів. Ренін-ангіотензинова система
Основним мінералокортикостероїдом є альдостерон (стероїд з альдегідною групою), який у 30-50 разів активніший за дезоксикортикостерон за впливом на мінеральний обмін. Альдостерону притаманна і глюкокортикоїдна активність, але у 5 разів менша, ніж активність кортизолу.
Синтез і секрецію альдостерону клітинами клубочкової зони стимулюють ангіотензин II, КТГ, простагландин Е, висока концентрація К+ і низька концентрація Nа+, гальмують дофамін і натрійуретичний фактор передсердя.
Мінералокортикоїди стимулюють збільшення реабсорбції Nа+,Cl і НСО-3 дистальними канальцями нирок і, одночасно, екскрецію К+. Затримка Nа+ і втрата К+ під дією альдостерону мають місце також у слинних і потових залозах, слизовій оболонці дистальних відділів товстого кишечника. Як і інші стероїдні гормони, альдостерон стимулює синтез у клітинах-мішенях невеликої кількості мРНК і, відповідно, білків. Вірогідно, деякі з цих білків сприяють перебудові мембрани і підвищенню ефективності роботи мембранних переносників Nа+ (пермеази). Альдостерон індукує також синтез мітохондріальних ферментів, що забезпечує енергією посилений транспорт Nа+, і Nа+,К+-АТФази, яка відкачує іони натрію з епітеліальних клітин канальця в міжклітинну рідину. Таким чином, ефект альдостерону не проявляється негайно, а через певний час, досягаючи максимуму через декілька годин. Секреція іонів К+ в дистальних канальцях здійснюється шляхом обміну на іони натрію. Затримка Nа+ збільшує затримку води і, таким чином, відновлює об'єм позаклітинної рідини. Відзначимо, що при цьому концентрація іонів в міжклітинній рідині зміниться дуже мало.
Альдостерон об'єднують в єдину систему з реніном і ангіотензином. Ця система нагадує систему гіпофіз-периферична залоза, оскільки ангіотензин проявляє тропну дію на клітини клубочкової залози кори надниркових залоз, так само, як КТГ регулює активність клітин пучкової зони. Ренін –це фермент, який утворюється в юкстагломерулярних клітинах нирок у відповідь на зниження артеріального тиску чи об'єму крові. У плазмі крові він діє на ангіотензиноген – білок, який секретується печінкою. Ренін гідролізує один пептидний зв'язок у молекулі ангіотензиногену, при цьому відщеплюється пептид із 10 амінокислотних залишків – ангіотензин І (неактивний). Потім фермент карбоксипептидилпептидаза, що знаходиться переважно в легенях, а також і в інших частинах судинного русла, відщеплює від ангіотензину І дві амінокислоти з утворенням ангіотензину ІІ. Цей октапептид є однією з найбільш активних судинозвужувальних речовин. Він сильніше діє на гладку мускулатуру артерій і артеріол, ніж вен. Одночасно ангіотензин ІІ стимулює синтез і секрецію альдостерону і спричиняє відчуття спраги. Судинозвужувальний вплив ангіотензину на артеріоли і затримка Nа+ та води альдостероном у кінцевому результаті відновлюють артеріальний тиск і об'єм рідини в організмі до вихідного рівня. Таким чином, перестають діяти стимули, що спонукали виділення реніну. Крім того, секреція реніну гальмується за механізмом зворотного зв'язку альдостероном і ангіотензином ІІ. Неадекватно висока продукція і секреція реніну призводить до ниркової гіпертонії.
Фізіологічними антагоністами ангіотензину є натрійуретичні гормони пептидної природи, що синтезуються в передсерді (тип А), мозку (тип В), багатьох тканинах (тип С). Ці пептиди пригнічують секрецію альдостерону і реабсорбцію іонів Nа+, стимулюють діурез, розширюють судини, знижують артеріальний тиск.
Рецептором натрійуретичного гормону служить гуанілатциклаза плазматичної мембрани клітин-мішеней. Зв'язування гормону зумовлює активацію ферменту, синтез цГМФ, що опосередковує клітинну відповідь.
Механізм дії альдостерону
Якщо ушкоджена значна частина кори надниркових залоз (інфекційним чи автоімунним процесами), то інтенсивніше порушуються функції, пов'язані з альдостероном, ніж глюкокортикоїдами. Відзначаються втрата натрію з сечею і затримка калію, дегідратація організму, гіпотонія і порушення периферичного кровообігу, ацидоз. Гіперкаліємія і гіпонатріємія зумовлюють м'язову слабкість, брадикардію, аритмію. При недостатньому харчуванні спостерігаються гіпоглікемія, зниження вмісту глікогену в тканинах, зниження азоту в сечі. Типовою ознакою є гіперпігментація шкіри через меланоцитостимулювальну дію КТГ. При хворобі Аддісона має місце висока чутливість організму до шкідливої дії різних факторів (інфекційних збудників, хімічних речовин, фізичних травм тощо). Зустрічаються генетичні порушення функції кори надниркових залоз, коли не синтезуються ферменти, необхідні для нормального утворення кортикостероїдів.
Чоловічі статеві гормони
Основний гормон з андрогенною активністю – тестостерон. За хімічною природою – С19-стероїд.Утворюється він в інтерстиціальних клітинах сім'яників (клітинах Лейдіга). Синтезується тестостерон, як і інші стероїдні гормони, із холестерину.
Сім'яники виробляють і секретують також невелику кількість дигідротестостерону й естрогенів. У надниркових залозах утворюються такі андрогени, як адреностерон, андростендіон, дегідроізоандростерон. Синтез і секреція тестостерону клітинами Лейдіга стимулюється лютеїнізуючим гормоном гіпофіза через аденілатциклазну систему. В свою чергу андрогени регулюють секрецію гонадоліберину і гонадотропінів за механізмом негативного зворотного зв'язку.
Тестостерон, як і інші жиророзчинні гормони, транспортується в крові специфічним білком, що синтезується в печінці. Він також зв'язує і переносить естрогени. Понад 97 % тестостерону зв'язано з білком у крові і тільки менше 3 % припадає на вільну, біологічно активну форму гормону. Концентрація тестостерону в плазмі крові чоловіків складає 3-10 мкг/л, а у жінок – 0,2-0,8 мкг/л. У печінці здійснюються різні реакції метаболізму андрогенів з утворенням сполук з меншою андрогенною активністю чи взагалі без неї. Ці сполуки відносяться до 17-кетостероїдів і виводяться з сечею у вигляді кон'югатів із глюкуроновою та сірчаною кислотами. 17-кетостероїди, які утворюються з тестостерону, складають приблизно 30 % всіх 17-кетостероїдів сечі і відрізняються від 17-кетостероїдів, які утворюються із андрогенів кори надниркових залоз, відсутністю оксигрупи у С11.
Андрогени беруть участь в ембріогенезі, розвитку первинних статевих ознак, формуванні вторинних статевих ознак (розподіл волосся на тілі, тембр голосу, тип відкладання жиру в організмі), рості скелета і скелетної мускулатури. Тестостерон разом з фолікулостимулювальним гормоном гіпофіза підтримує сперматогенез. Андрогени мають механізм дії, спільний для стероїдних гормонів. Тестостерон у деяких клітинах-мішенях, зокрема простати, сім'яних міхурців, діє після перетворення в дигідротестостерон. На кістки, м'язи і нирки діє, вірогідно, сам тестостерон. Рецепторні білки андрогенів знаходяться у цитоплазмі і ядрі клітин-мішеней. Андрогени мають виражену анаболічну дію, стимулюють синтез білків, нуклеїнових кислот, фосфоліпідів мембран. Серед білків, синтез яких прискорюється тестостероном, відомі РНК-полімерази, рецепторний білок, білки рибосом. Андрогени затримують азот, кальцій і фосфор в організмі, збільшують загальну масу скелетних м'язів. Синтезовані аналоги андрогенів, у яких анаболічна дія у 20 разів більша, ніж у тестостерону, а андрогенна активність менша. Такі речовини широко використовуються спортсменами, хоч вони можуть спричинити ураження печінки, зокрема виникнення пухлин.
У клітинах ЦНС тестостерон перетворюється в естрогени, які беруть участь у регуляції секреції гіпофізарних гонадотропінів і статевої поведінки. В організмі здорової жінки синтезуються і секретуються тестостерон і його попередники, але значно менше, ніж у чоловіків (в середньому 250 мкг/добу проти 7000 мкг/добу). Крім того, в надниркових залозах і яєчнику утворюється менш активний андростендіон.
При підвищеній продукції андрогенів в організмі жінок виникають різні прояви вірилізму (маскулінізації).
Тестостерон і його синтетичні аналоги використовуються для лікування раку молочної залози в жінок.
Жіночі статеві гормони
До жіночих статевих гормонів відносяться естрогени і прогестерон. Естрогени утворюються в яєчниках. За хімічною природою – С18-стероїди із ароматичним кільцем А.
Естрогени
Основний, найбільш активний, естроген, бета-естрадіол, секретується фолікулами яєчників. Естрон і естріол утворюються в основному при метаболізмі естрадіолу в печінці і плаценті. Естріол також синтезується плацентою із стероїдного попередника (дегідроепіандростерону), який надходить із надниркових залоз плоду.
Естрадіол синтезується, як і всі стероїдні гормони, з холестерину, причому проміжним продуктом є тестостерон. Таким чином, естрогени утворюються з чоловічого статевого гормону – тестостерону. Заключною стадією синтезу є ароматизація першого кільця; вона включає три реакції гідроксилювання за участю О2, НАДФН і цитохрому Р-450. Синтез і секреція естрогенів стимулюються лютеїнізуючим і фолікулостимулювальним гормонами гіпофіза через аденілатциклазну систему.
Естрогени гальмують за механізмом негативного зворотного зв'язку секрецію гонадоліберину гіпоталамуса і гонадотропінів гіпофіза. У печінці естрогени і їх метаболіти утворюють кон'югати із глюкуроновою або сірчаною кислотами – неактивні сполуки, що виводяться з сечею. Естрогени відповідають за ріст і розвиток органів репродуктивної системи у процесі статевого дозрівання жінки та здатність до розмноження у репродуктивному періоді. У період дитинства естрогени секретуються у настільки малій кількості, що не індукують розвитку репродуктивних органів. Причиною низької секреції естрогенів є загальмованість секреції гонадотропних гормонів на рівні ЦНС. У відповідь на дію гонадотропінів підвищена продукція естрогенів приводить до прискорення росту і розвитку матки, піхви, зовнішніх статевих органів, таза, молочних залоз. У цілому естрогени діють як гормони росту на ті тканини, які прямо чи опосередковано зв'язані з процесом розмноження. У дорослих жінок на протязі репродуктивного періоду продукція естрогенів зазнає циклічних змін.
Молекулярний механізм дії естрогенів на клітини-мішені такий, як у всіх стероїдних гормонів. Вони проникають у цитоплазму, з'єднуються з рецепторами, комплекс гормон-рецептор переміщається в ядро і взаємодіє із певними ділянками ДНК. В результаті індукується синтез певних мРНК і, відповідно, специфічних білків. Один із ферментів, активність якого значно збільшується, під дією естрогенів, є орнітиндекарбоксилаза. Вона каталізує утворення путресцину, субстрату для біосинтезу поліамінів, необхідних для проліферації тканин.
Анаболічна дія естрогенів значно слабша, ніж андрогенів. Естрогени впливають на мінеральний обмін у кістках, гальмують резорбцію кісток, про що свідчить розвиток остеопорозу в період після менопаузи. Вважають, що естрогени діють на мінеральний обмін опосередковано через кальцитонін і вітаміни D. Естрогени впливають на синтез білків у печінці, зокрема синтез білків – переносників гормонів, факторів згортання крові, ангіотензиногену, білків, що входять до складу ліпопротеїнів високої густини.
Естрогени малоактивні при пероральному надходженні, оскільки швидко інактивуються в печінці. Такі синтетичні аналоги, як діетилстильбестрол і гексестрол, мають високу активність при пероральному надходженні і широко застосовуються у клінічній практиці. За хімічною структурою вони не є стероїдами, але їх просторова структура така, що вони взаємодіють із рецепторами естрогенів.
Прогестерон
Прогестерон утворюється в жовтому тілі, плаценті і надниркових залозах із холестерину. Клітини жовтого тіла і плаценти продукують і естрогени, але основний їх продукт – прогестерон. На відміну від естрогенів, прогестерон має 21 атом вуглецю, як кортикостероїди.
Синтез прогестерону жовтим тілом стимулюється лютеїнізуючим гормоном гіпофіза, а на ранніх стадіях вагітності – хоріонічним (плацентарним) гонадотропіном. Інактивується прогестерон у печінці шляхом відновлення до прегнандіолу, який далі утворює кон'югати із глюкуроновою чи сірчаною кислотами, що виділяються із сечею.
Секреція прогестерону жовтим тілом готує ендометрій матки для імплантації заплідненої яйцеклітини. В період вагітності прогестерон проявляє гальмуючий вплив на мускулатуру матки, "заспокоюючи" її скорочення. Вплив цей пов’язаний із зміною вибіркової проникності мембран гладком’язових клітин для натрію і калію. Введення прогестерону призводить до зниження рівня калію і підвищення рівня натрію в міометрії. Разом з естрогенами прогестерон стимулює ріст молочних залоз. Естрогени стимулюють ріст системи проток, а прогестерон – ріст нових залозистих елементів залози. Естрогени і прогестерон перешкоджають початку лактації під час вагітності, блокуючи дію пролактину на молочну залозу.
Гормональний контроль статевого циклу
Цикл зводить в єдине ціле різні процеси: дозрівання яйцеклітини і її овуляцію, ріст, розвиток і інволюцію жовтого тіла, підготовку ендометрія матки до імплантації ембріона, активність структур мозку і гіпофіза, що контролюють співвідношення гонадотропних гормонів. Координація цих процесів досягається узгодженими в часі коливаннями швидкостей продукції і секреції ряду гормонів.
Ритмічні цикли, які починаються після статевого дозрівання, зв'язані із секрецією гонадотропних гормонів гіпофіза, які в період до статевої зрілості не секретуються.
Вважають, що початок статевого дозрівання запускаються зміною гормональної активності епіфіза (шишкоподібної залози), яка продукує гормон мелатонін – універсальний регулятор біологічних ритмів. За структурою мелатонін є похідним триптофану.
Продукція його стимулюється у темноті і характеризується циркадним ритмом. У тварин мелатонін впливає на пігментацію шкіри, викликаючи її освітлення (протилежна дія до МСГ).
В організмі жінок мелатонін гальмує секрецію гонадотропінів двома шляхами:
1) впливає на клітини гіпоталамуса, пригнічуючи вивільнення гонадоліберину;
2) у гіпофізі зменшує стимулювальну дію гонадоліберину на продукцію і секрецію гонадотропних гормонів. На початку статевого дозрівання епіфіз знижує секрецію мелатоніну, що запускає утворення гіпоталамусом гонадоліберину, який, в свою чергу, стимулює секрецію ФСГ і ЛГ гіпофіза. Циркадний ритм звільнення гонадотропінів, естрогенів і прогестерону є наслідком ритму секреції мелатоніну епіфізом.
У перші дні менструальної кровотечі збільшується концентрація ФСГ у крові. Розвивається декілька фолікулів, але тільки один дозріває.
При дозріванні клітин внутрішнього шару фолікула виробляється все більше естрогенів. За механізмом негативного звязку естрогени гальмують секрецію ФСГ. Секреція лютеїнізуючого гормону в цей час підвищується. Перед самою овуляцією зростає під дією ЛГ секреція естрогенів і прогестерону фолікулом, що служить сигналом для різкого підйому секреції лютеїнізуючого гормону гіпофізом. Тут діє механізм позитивного зворотного звязку. Гонадотропні гормони стимулюють перетворення у фолікулярній рідині плазміногену в плазмін. Він зумовлює ряд перетворень, що завершуються розривом фолікула і звільненням яйцеклітини. На цей процес впливають також прогестерон і простагландин F2. Фолікул перетворюється у жовте тіло: заповнюється лютеальними клітинами, що мають жовте забарвлення і багаті ліпідами. Під впливом ЛГ клітини жовтого тіла синтезують все більше прогестерону і естрадіолу. У результаті їх дії на гіпоталамус гальмується секреція гіпофізарних гонадотропінів. Прогестерон впливає на ендометрій матки, яка функціонально готується до прийому заплідненої яйцеклітини.
Якщо вагітність не настає, жовте тіло дегенерує, внаслідок чого різко знижується концентрація прогестерону і естрогенів, зростає рівень ФСГ, що запускає нову хвилю дозрівання фолікулів. В ендометрії виникають геморагічні і дегенеративні зміни: кровотеча, відторгнення поверхневих шарів ендометрія, слизу, що усуваються із менструальною кров'ю.
Якщо запліднена яйцеклітина імплантується в ендометрій, то жовте тіло продовжує продукувати прогестерон і естрогени на ранніх стадіях вагітності, а далі вони утворюються в плаценті. Крім того, плацента продукує хоріонічний гонадотропін, плацентарний лактоген (соматомамотропін), плацентарні кортикотропін і тиреотропін. Плацента і плід тісно пов'язані між собою, так що синтез естріолу відбувається за участю материнського організму, плаценти і плода. Холестерин із печінки матері надходить у плаценту, перетворюється у прегнелон, який в надниркових залозах плода перетворюється в один із андрогенів (дегідроепіандростерон), а із нього у плаценті синтезується естріол. Екскреція естріолу із сечею матері може служити показником життєздатності плода.
Під час вагітності естрогени і прогестерон необхідні для розвитку і росту матки і молочних залоз. Естрогени стимулюють ріст гладком'язових клітин матки, збільшують її м'язову масу, забезпечуючи тим самим скоротливу активність матки під час пологів. Прогестерон перешкоджає ефективним координованим скороченням матки, перетворюючи їх в окремі слабкі фібриляції. Стимул, який запускає пологи, точно не визначений. Відомо, що на початку пологів відбувається викид кортизолу самим плодом і синтез простагландинів, припинення дії прогестерону, секреція нейрогіпофізом окситоцину і зростання кількості рецепторів до нього на клітинах міометрія.
Класифікація та номенклатура вітамінів
За фізико-хімічними властивостями та відношенням до обміну речовин вітаміни поділяють на дві групи: вітаміни, розчинні у воді, і вітаміни, розчинні у жирах. Водорозчинні вітаміни безпосередньо беруть участь в обміні речовин як коферменти або складові компоненти коферментів. Жиророзчинні вітаміни не входять до складу ферментів і впливають на обмін речовин опосередковано, створюючи умови для оптимальної дії ферментів на мембранних структурах. Вони виконують роль модуляторів структури і функцій мембран. У зв'язку з цим, жиророзчинні вітаміни в організмі виконують ще антимутагенну функцію, захищаючи генний апарат від пошкоджень хімічними та фізичними факторами. Це зв'язано із вираженими антиоксидантними властивостями жиророзчинних вітамінів: вони здатні знешкоджувати активні форми кисню та вільні радикали й гальмувати процеси пероксидного окиснення біополімерів (нуклеїнових кислот, білків, ліпопротеїнових комплексів).
Вітаміни, розчинні у жирах
1. Вітамін А, ретинол, ретиноєва кислота, антиксерофтальмічний.
2. Вітамін D, антирахітний.
3. Вітамін Е, токофероли, антистерильний, вітамін розмноження, антиоксидант.
4. Вітамін К, нафтохінони, антигеморагічний.
5. Вітамін F, есенціальні жирні кислоти, антисклеротичний.
Вітаміни, розчинні у воді
1. Вітамін В1, тіамін, антиневритний.
2. Вітамін В2, рибофлавін, вітамін росту.
3. Вітамін В3, пантотенова кислота, антидерматитний.
4. Вітамін В5, РР, нікотинамід, нікотинова кислота, антипелагричний.
5. Вітамін В6, піридоксин, піридоксамін, піридоксаль, антидерматитний.
6. Вітамін В10, Вс, фолієва кислота, фоліацин, фактор росту, антианемічний.
7. Вітамін В12, ціанкобаламін, антианемічний.
8. Вітамін Н, біотин, антисеборейний.
9. Вітамін С, аскорбінова кислота, антискорбутний.
10. Вітамін Р, біофлавоноїди, фактор проникності, капілярозміцнюючий.
Вітаміноподібні речовини
Убіхінон, кофермент Q.
В4, холін, фосфохолін.
В8, інозит.
N, ліпоєва кислота.
Вт, карнітин.
В13, оротова кислота, фактор росту.
В15, пангамова кислота, антианоксичний.
U, S-метилметіонін, антивиразковий.
ПАБК, параамінобензойна кислота, вітамін для росту мікроорганізмів.
Відокремлення від вітамінів групи вітаміноподібних речовин часто умовне. Останні за біологічними функціями подібні до вітамінів, але потрібні в значно більших кількостях.
Водорозчинні вітаміни
До водорозчинних відносяться вітаміни групи В та інші вітаміни (С, Р, Н) і вітаміноподібні водорозчинні сполуки. На противагу жиророзчинним вітамінам, які відіграють у клітинах роль модуляторів клітинних мембран, водорозчинні вітаміни є основними коферментами або входять до складу коферментів різних ферментних систем. Водорозчинні вітаміни в тканинах зв'язані з білками, не мають провітамінів і не викликають гіпервітамінозів. Розглянемо вітаміни групи В.
Вітамін В1
Вітамін В1 – тіамін, антиневритний фактор. Добова потреба в тіаміні для дорослої людини складає 1-3 мг. Міститься він в хлібі грубого помолу, горосі, квасолі, а також тваринних продуктах: печінці, м'ясі та інших. Всмоктується в тонкому кишечнику і кров'ю заноситься в печінку, де під впливом ферменту тіамінфосфокінази фосфорилюється до тіамінфосфату, тіаміндифосфату та тіамінтрифосфату. Найбільш вивчена роль тіаміндифосфату. Із печінки тіамін розноситься в різні органи. Найбільше його є в м'язах (до 50 %), а решта – в печінці та інших органах і тканинах.
Тіаміндифосфат (кокарбоксилаза) входить як кофермент до складу піруватдегідрогенази, альфа-кетоглутаратдегідрогенази і транскетолази. Завдяки цьому бере участь в окисненні пірувату й альфа-кетоглутарату в мітохондріях, а отже, у вилученні енергії з продуктів розщеплення вуглеводів, білків та жирів. Транскетолаза забезпечує перебіг неокиснювальної фази пентозофосфатного циклу, який призводить до нагромадження НАДФН2 і рибозо-5-фосфату. За рахунок цього вітамін В1 необхідний для синтезу жирних кислот, холестерину, стероїдних гормонів, знешкодження токсичних речовин, ліків тощо. А рибозо-5-фосфат використовується для синтезу нуклеїнових кислот, нуклеотидних коферментів, нуклеотидів. У мозковій тканині знаходиться в достатній кількості тіамінтрифосфат, що має відношення до синаптичної передачі нервових імпульсів.
Гіповітаміноз В1 характеризується зниженням вмісту названих вище коферментів у тканинах і, як наслідок, послабленням активності транскетолази та дегідрогеназ кетокислот. Саме тому вміст піровиноградної та альфа-кетоглутарової кислот підвищується в крові й сечі, зменшується використання їх в енергозабезпеченні тканин, насамперед мозку (мозкова тканина живиться переважно вуглеводами). Через нестачу НАДФН2 і рибозо-5-фосфату, що викликано пригніченням транскетолазної реакції, загальмовуються процеси біосинтезу, зокрема замінних амінокислот. Тому в обміні речовин процеси катаболізму будуть переважати над анаболізмом.
Через нестачу вітаміну В1 в організмі розвивається ряд патологічних проявів, які називаються бері-бері. Зміни спостерігаються з боку метаболізму і функцій органів травлення, серцево-судинної, нервової та м'язової систем. Розлади шлунково-кишкового тракту проявляються у вигляді різкого зменшення апетиту, зниження секреції шлункового соку і соляної кислоти, атонії кишечника, в'ялих закрепів. Характерними рисами бері-бері є різка атрофія м'язової тканини, зниження скоротливої здатності скелетних м'язів (виражена м'язова слабкість)
Рис. Авітаміноз В1
Спостерігаються зменшення сили серцевих скорочень, розширення правого шлуночка, тахікардія і гостра серцево-судинна недостатність. Неврологічними ознаками недостачі вітаміну В1 найчастіше є такі зміни: поступове зниження периферичної чутливості, втрата деяких периферичних рефлексів, сильний біль по ходу нервів, корчі, розлади вищої нервової діяльності (страх, зниження інтелекту, галюцинації). Але зараз вважається, що бері-бері в країнах, де харчуються полірованим рисом, не є проявом чистого гіповітамінозу В1. Більш правдоподібним є погляд, що бері-бері є наслідком нестачі в продуктах харчування цілого ряду вітамінів (поліавітаміноз), зокрема В1, В2, РР, С та інших. У Європі гіповітаміноз описаний у вигляді енцефалопатій (синдром Верніке) або ураження серцево-судинної системи (синдром Вейса). При гіповітамінозі В1 швидше виникають розлади в шлунково-кишковій системі й порушення психіки, настають зміни серцево-судинної і м'язової систем. Дещо пізніше розвиваються ураження периферичної нервової системи (розлади чутливості, біль по ходу нервів та інші). Ці зміни завершуються контрактурами, атрофією та паралічами нижніх, а потім і верхніх кінцівок.
Окремою формою тіамінової недостатності є уроджені порушення метаболізму вітаміну, наприклад тіамінзалежна анемія.
У медицині використовуються тіамін і тіаміндифосфат (кокарбоксилаза) для покращання засвоєння вуглеводів, у лікуванні цукрового діабету, гіповітамінозів, дистрофій міокарда, уражень м'язів та центральної і периферичної нервової системи.
Вітамін В2 (рибофлавін)
Рибофлавін складається з трициклічної сполуки ізоалоксазину та спирту рибітолу (похідного рибози), звідки і походить його назва. Надходить в організм із продуктами харчування, частково синтезується кишковими мікроорганізмами. Найбільше вітаміну В2 міститься в печінці, нирках, домашньому сирі, жовтку курячого яйця. Добова потреба його для дорослої людини становить 1-3 мг. Всмоктується рибофлавін у тонкому кишечнику за допомогою простої дифузії. У тканинах організму він перетворюється на ФМН і ФАД.
ФМН і ФАД як коферменти входять до складу флавінових ферментів, що беруть участь у багатьох окиснювальних реакціях клітин: передачі електронів і протонів у дихальному ланцюзі, окисненні пірувату, альфа-кетоглутарату, жирних кислот, біогенних амінів, альдегідів тощо.
Гіповітаміноз В2 характеризується зниженням вмісту коферментів, насамперед ФМН, що проявляється пригніченням процесів тканинного дихання і спричиняє затримку росту, посилений розпад тканинних білків.
Досить специфічними симптомами для гіповітамінозу В2 є ураження епітелію слизових, шкіри і рогівки ока: сухість слизових губ, порожнини рота. Слизова рота яскраво-червоного кольору, в його кутиках і на губах тріщини, лущення шкіри на обличчі, сухість кон'юнктиви, її запалення, проростання судинної сітки в рогівку. Васкуляризація рогівки полегшує надходження кисню в її центральну безсудинну зону та є проявом компенсації функції рогівки, викликаної дефіцитом флавінових коферментів, необхідних для окисно-відновних процесів.
Сухість кон'юнктиви, її запалення, проростання судинної сітки в рогівку при нестачі вітаміну В2
Практичне використання. У медицині рибофлавін застосовується при гіповітамінозі В2, а також для лікування захворювань шкіри, очей, спричинених іншими чинниками, для прискорення загоювання ран, виразок, а також для лікування уражень печінки, при отруєнні чадним газом тощо.
Вітамін В3 (пантотенова кислота, антидерматитний)
Джерелом пантотенової кислоти для людини є кишкові мікроорганізми і продукти харчування. Найбільше її міститься в дріжджах, печінці, курячих яйцях, молоці, м'ясі, стручкових тощо. Добова потреба у вітаміні В3 для дорослої людини складає приблизно 10 мг.
Біологічна функція пантотенової кислоти: вона входить до складу коферменту А (кофермент ацилювання). Кофермент А утворюється в результаті приєднання до СООН-групи бета-аланіну пантотенової кислоти залишку тіоетиламіну та залишку АДФ до гамма-ОН-групи масляної кислоти.
Функціонально активною групою коензиму А (КоА) є кінцева сульфгідрильна група, яка може зазнавати ацилювання з утворенням ацил-КоА або знаходитися в деацильованому стані КоА-SH. Крім того, вітамін В3 входить до складу фосфопантотеїну, що є коферментом ацилпереносного білка синтетази жирних кислот.
Практичне застосування. У медицині використовують пантотенат кальцію і КоА для лікування захворювань шкіри, уражень печінки, міокардіодистрофій, а також у парфумерії.
Вітамін В5 (РР, нікотинамід, протипелагричний)
Нікотинова кислота досить поширена в рослинних і тваринних продуктах. Для людини основним її джерелом є хліб, картопля, рис, м'ясо, печінка, нирки, морква та інші продукти. Добова потреба в ніацині для дорослої людини складає приблизно 25 мг.
Всмоктується вітамін РР у тонкій кишці простою дифузією. З ентероцитів він потрапляє в кров, якою переноситься в печінку та інші органи. У клітинах нікотинова кислота перетворюється в НАД+ і НАДФ+.
Біологічні функції. У складі НАД+ і НАДФ+ ніацин бере участь в обміні речовин. Є приблизно сотня нікотинамідзалежних ферментів. НАД+ і НАДФ+ є коферментами багатьох дегідрогеназ, необхідних для вироблення енергії в клітині: виступають акцепторами і проміжними переносниками атомів водню на початкових стадіях окиснення вуглеводів, жирних кислот, амінокислот, гліцерину, на стадії циклу Кребса і в термінальних стадіях дегідрування в дихальному ланцюзі та монооксигеназному ланцюзі.
Таким чином, вітамін В5 бере участь в енергозабезпеченні клітин і в знешкодженні шляхом окиснення природних та чужорідних речовин (монооксигеназний ланцюг окиснення).
НАДФН2 як донор атомів водню використовується в біосинтетичних відновних реакціях (синтез жирних кислот, холестерину, стероїдних гормонів та ін.).
Крім того, НАД+ є субстратом для реакції синтезу полі-АДФ-рибози. Цей процес має відношення до регуляції утворення нуклеїнових кислот у клітинному ядрі. НАД+ і НАДФ+ виступають також у ролі алостеричних ефекторів ферментів енергетичного обміну. Так, НАДН є алостеричним інгібітором ферментів циклу Кребса (ізоцитратдегідрогенази, малатдегідрогенази та ін.). Надлишок НАДФН викликає інгібування глюкозо-6-фосфатдегідрогенази.
Недостатність ніацину проявляється захворюванням, що називається пелагрою. Важливо зазначити, що гіповітаміноз В5 супроводжується одночасним гіповітамінозом В2 та В6. Це пов'язано з тим, що для утворення нікотинової кислоти з триптофану потрібні коферменти рибофлавіну та піридоксину.

Дитина, хвора на пелагру
Частими проявами гіповітамінозу є також порушення інтелекту (деменція) і функції периферичних нервів (неврити).
Практичне застосування. У медицині нікотинову кислоту та її амід використовують для лікування пелагри, дерматитів, викликаних іншими чинниками, невритів, дистрофій серцевого м'яза, а також у клініці як судинорозширювальний засіб.
Вітамін В6 (піридоксин, антидерматитний)
Вітамін В6 у великій кількості міститься в продуктах рослинного і тваринного походження. Для людини джерелом вітаміну В6 є кишкові бактерії, а також хліб, горох, квасоля, картопля, м'ясо, печінка, нирки й ін.; менше його в капусті й моркві. Добова потреба у вітаміні В6 для дорослої людини складає в середньому 2-3 мг.
Всмоктується вітамін у тонкій кишці простою дифузією. З ентероцитів він потрапляє в кров, а звідси переноситься до різних тканин, де під впливом специфічних кіназ піридоксаль і піридоксамін фосфорилюються і перетворюються в піридоксальфосфат та піридоксамінфосфат.
Біологічні функції. Біологічна роль вітаміну В6 пов'язана з коферментами (піридоксальфосфатом та піридоксамінфосфатом), які знаходяться в усіх клітинах організму. З участю вітаміну В6 у складі коферменту піридоксальфосфату відбуваються реакції трансамінування амінокислот та окиснення біогенних амінів, декарбоксилювання амінокислот та їх ізомеризація, синтез нікотинаміду з триптофану, біосинтез гему, гамма-аміномасляної кислоти й ін. Піридоксальфосфат входить до складу майже всіх класів ферментів. Звідси зрозуміло, що при нестачі вітаміну В6 можуть спостерігатися численні порушення обміну речовин, зокрема амінокислот.
Гіповітаміноз В6
Піридоксинова недостатність може спостерігатися в дітей і проявлятися підвищеною збудливістю центральної нервової системи і періодичними судомами. Причиною таких змін є недостатня кількість гальмівного медіатора центральної нервової системи гамма-аміномасляної кислоти (ГАМК), що утворюється з глутамінової кислоти під впливом ферменту декарбоксилази з участю коферменту піридоксальфосфату. В дорослих людей гіповітаміноз може виникати при тривалому лікуванні туберкульозу протитуберкульозним препаратом ізоніазидом, який за своєю природою є антагоністом (антивітаміном) піридоксалю. У цих хворих також підвищена збудливість центральної нервовї системи, виникають поліневрити і спостерігаються ураження шкіри, характерні для гіповітамінозу РР.
Вітамін В10 (фолієва кислота, фоліацин, антианемічний)
Для дорослої людини добова потреба в цьому вітаміні складає 0,2‑0,5 мг. Основним джерелом фолієвої кислоти для людини є рослинна і тваринна їжа. Багато її міститься в листках рослин, овочах, фруктах, а також у печінці та м'ясі. Додатковим джерелом фолієвої кислоти є і мікрофлора кишечника. В організмі дорослої людини міститься 7-12 мг цієї кислоти, з них більше половини – в печінці, нирках і слизовій кишечника.
Всмоктується фолієва кислота в тонкій кишці, де з неї утворюється тетрагідрофолієва кислота (ТГФК). Процес відбувається у дві стадії. Спочатку за допомогою ферменту фолатредуктази утворюється дигідрофолієва кислота (ДГФК), яка з участю іншого ферменту – дигідрофолатредуктази – відновлюєтьсяя до ТГФК:
Гіповітаміноз В10
При нестачі фолієвої кислоти порушується обмін одновуглецевих груп, що проявляється у вигляді мегалобластичної анемії. Спостерігаються зменшення кількості еритроцитів, зниження вмісту гемоглобіну в периферичній крові, з’являються великі клітини (мегалобласти), настає зменшення кількості лейкоцитів (лейкопенія). Мегалобластична анемія найчастіше є результатом нестачі вітаміну В10 чи В12 або одночасно обох. Нестача цих вітамінів відбивається, насамперед, на біосинтезі ДНК у кровотворних клітинах, що зазнають швидкого поділу і проліферації (не утворюються тимідилова кислота, пуринові нуклеотиди, порушується метилювання нуклеїнових кислот і, як наслідок, змінюється еритропоез).
Практичне застосування. У практичній медицині фолієву кислоту використовують для лікування мегалобластичної анемії, отруєнь метиловим спиртом (включає в обмін формільний радикал), для стимуляції проліферації клітин.
Вітамін В12 (ціанокобаламін, антианемічний)
Вітамін В12 не синтезується ні в рослинах, ні в тваринних організмах. Його здатні утворювати тільки мікроорганізми.
Основним джерелом вітаміну В12 для людини є продукти тваринного походження – печінка, нирки, серце, м'ясо та ін. Частково він синтезується мікрофлорою кишечника за умов надходження з їжею кобальту. Добова потреба у вітаміні В12 для дорослої людини складає 2,5-5 мкг.
Всмоктування кобаламіну відбувається в тонкій кишці. Але для цього необхідний так званий внутрішній фактор Кастла, що утворюється в обкладкових клітинах шлунка. За своєю природою він являє собою глікопротеїн, який має здатність вибірково зв'язуватись з вітаміном В12. Комплекс вітамін В12 – внутрішній фактор заноситься в кишечник, тут відбувається приєднання його до специфічних рецепторів мембран ентероцитів, перенесення через мембрану і всмоктування. При потраплянні кобаламіну в кров глікопротеїн відщеплюється від комплексу. У крові кобаламін переноситься спеціальними транспортними білками (транскобаламіном І і транскобаламіном ІІ), що відносяться до альфа- і бета-глобулінів.
Біологічні функції. Вітамін В12 бере участь у багатьох хімічних перетвореннях, але механізм їх ще недостатньо вивчений. Встановлено, що кофермент метилкобаламін входить до складу ферменту, який переносить метильну групу 5-метилтетрагідрофолієвої кислоти на гомоцистеїн з утворенням метіоніну:
![]()
Таким чином, вітамін В12 функціонально тісно пов'язаний із фолієвою кислотою. Разом з нею він сприяє синтезу метіоніну, утворенню креатину, пуринових і піримідинових основ, амінокислот, білків, нуклеїнових кислот і ін. Кофермент дезоксиаденозилкобаламін у складі ферменту метилмалоніл КоА-мутази перетворює метилмалоніл-КоА в сукциніл-КоА. Ця реакція має місце в завершальній стадії окиснення жирних кислот з непарною кількістю вуглецевих атомів у циклі Кребса, окисненні бокового ланцюга холестерину, тиміну, амінокислот (метіонін, ізолейцин, треонін, валін). Встановлено, що кобаламін сприяє депонуванню та утворенню коферментних форм фолієвої кислоти і за рахунок цього бере участь в синтезі ДНК і проліферації кровотворних клітин.
Гіповітаміноз В12 виникає найчастіше як ускладнення гастриту, зокрема при гіпо- або анацидних гастритах, після оперативного видалення шлунка чи частини його, де виробляється внутрішній фактор. Тому вітамін В12 не всмоктується, він виводиться з калом і як наслідок розвивається злоякісна анемія Адісона-Бірмера, або мегалобластична. Порушення кровотворення при цьому гіповітамінозі аналогічні тим, що мали місце у хворих з дефіцитом фолієвої кислоти. Але при нестачі кобаламіну спостерігається підвищення виділення з сечею метилмалонової кислоти, яка не засвоюється; можливі також ураження задніх і бокових стовпів спинного мозку (фунікулярний мієлоз).
Практичне застосування. Кобаламіни застосовуються для лікування мегалобластичної анемії, уражень спинного мозку і периферичних нервів. Найдоцільніше використовувати кобаламіни в поєднанні з фолієвою кислотою і залізом, позаяк вони необхідні для синтезу гемоглобіну в кровотворних органах.
Вітамін С (аскорбінова кислота, антискорбутний)
Середньодобова потреба людини у вітаміні С складає 80-100 мг. Основним його джерелом є овочі, фрукти, ягоди. Найбільше аскорбінової кислоти міститься в шипшині, чорній смородині; з овочів багаті нею капуста, помідори, червоний перець, картопля і ін.
Всмоктується вона у всьому шлунково-кишковому тракті, але найкраще в тонкій кишці простою дифузією. У тканинах аскорбінова кислота зв'язується з білками. Вона буває вільною і вступає в окисно-відновні реакції. Найбільшу кількість аскорбінової кислоти виявлено в печінці, надниркових залозах, легенях.
У процесі метаболізму аскорбінова кислота перетворюється в дегідроаскорбінову, що спричиняється різними чинниками, зокрема киснем, метиленовою синькою, перекисом водню та ін. Цей процес не супроводжується зниженням вітамінної активності. Але дегідроаскорбінова кислота є не дуже стійкою сполукою і тому в слабколужному або нейтральному середовищі перетворюється в дикетогулонову кислоту, що не проявляє вітамінних властивостей. Біологічну роль аскорбінової кислоти найчастіше пов'язують з участю в окисно-відновних процесах. Але ферментів, у яких кислота відігравала б роль коферменту, досі не виявлено. Встановлено, що вона служить донором водню для відновлення різних біологічних субстратів. Зокрема відомо, що аскорбінова кислота може відновлювати дисульфідні зв'язки до сульфідних груп, активуючи цим самим ряд ферментів. В свою чергу дегідроаскорбінова кислота може ферментативно відновлюватись у тканинах організму за участю глутатіону. Біологічна роль аскорбінової кислоти тісно пов'язана з обміном білків, вуглеводів, мінеральних речовин. За участю аскорбінової кислоти перебігають процеси гідроксилювання з утворенням цілого ряду біологічно активних речовин. Так, через гідроксилювання триптофан перетворюється в 5-гідрокситриптофан, який служить основою для утворення медіатора серотоніну. Аскорбінова кислота необхідна для процесів гідроксилюванняя під час перетворення холестерину в стероїдні гормони, для перетворення 3,4-дигідроксифенілетиламіну в норадреналін. Вона сприяє звільненню заліза із феритину та трансферину, що забезпечує проникнення його в тканини. В кишечнику за участю аскорбінової кислоти відбувається відновлення Fe3+ до Fe2+, що необхідно для його всмоктування. Відновлення фолієвої кислоти до коферментної форми ТГФК також відбувається за участю аскорбінової кислоти. Без аскорбінової кислоти не відбувається процес гідроксилювання проліну і лізину, а отже, і перетворення проколагену в колаген, який є головним позаклітинним компонентом сполучної тканини.
Гіповітаміноз С
Завдяки участі аскорбінової кислоти в багатьох біохімічних процесах при обмеженому її надходженні розвивається ряд метаболічних порушень та клінічних проявів. Нестача в організмі аскорбінової кислоти відома під назвою цинги, або скорбуту, яка в минулі століття нерідко мала характер епідемій (рис.).
Рис. Виражений стоматит при С авітамінозі
Найхарактернішою ознакою гіповітамінозу С є втрата організмом здатності виробляти основну міжклітинну "цементуючу" речовину – колаген. Порушується також синтез глікопротеїнів (мукополісахаридів), що призводить до ураження судинних стінок і опорних тканин. Зміни спостерігаються в першу чергу в кровоносній системі: судини стають ламкими і проникними, що супроводжується дрібними крововиливами під шкіру і в шкіру – так звані петехії. Можливі крововиливи в слизові оболонки та внутрішні органи. Одночасно з'являється кровоточивість із ясен, деполімеризація і руйнування одонтобластів і остеобластів, що призводить до розхитування та випадання зубів.
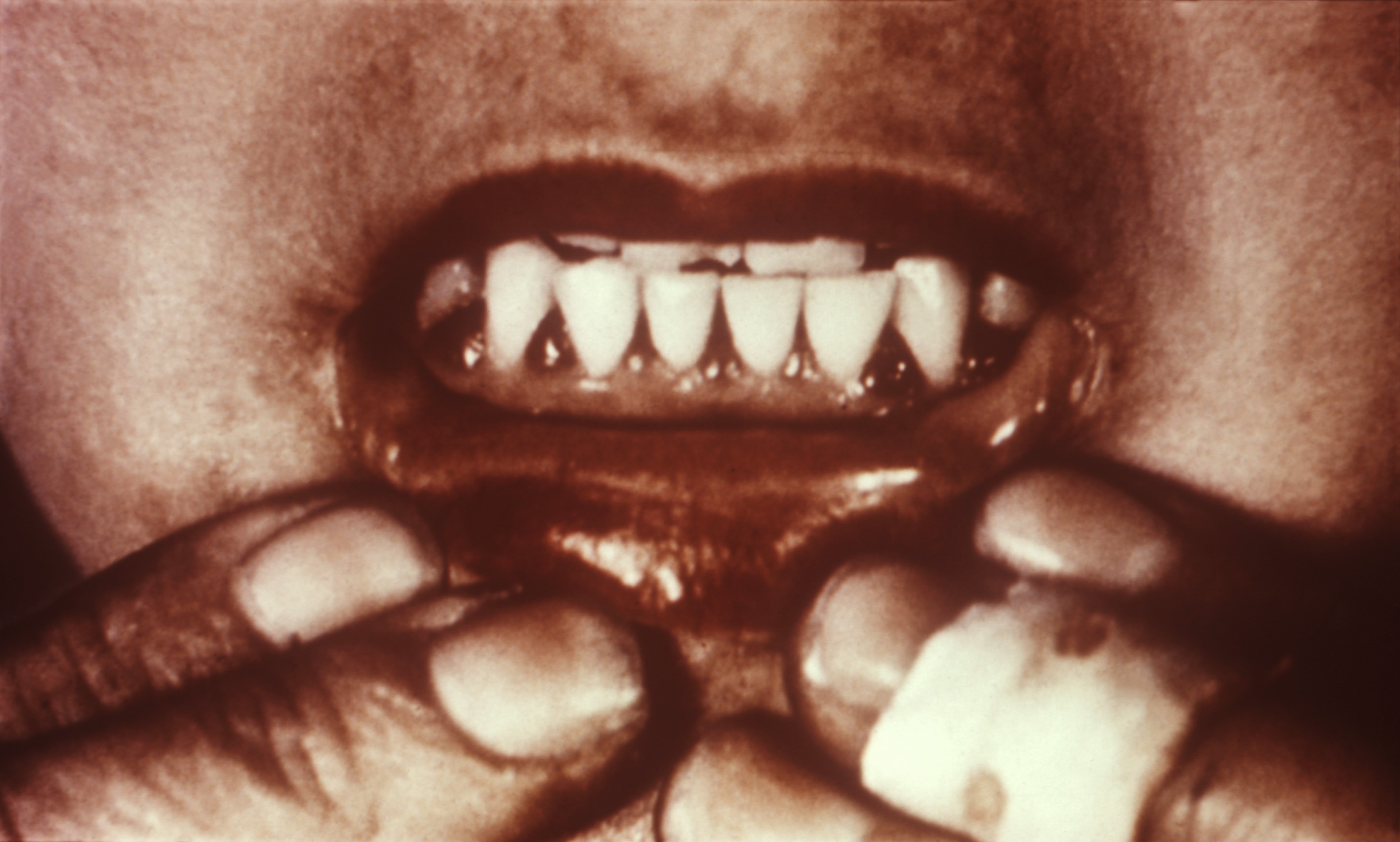
Рис. Ураження десен при скорбуті
У хворих на цингу, крім цього, спостерігаються набряки нижніх кінцівок, болі під час ходьби, болі в серці, серцебиття, задишка, загальна слабкість, схуднення. Як ускладнення гіповітамінозу С у хворих може розвинутись анемія – не використовується залізо і фолієва кислота в процесі гемопоезу.
Практичне застосування. В медицині аскорбінову кислоту застосовують для стимуляції регенеративних процесів, ураження сполучної тканини і, звичайно, для профілактики та лікування цинги.
Жиророзчинні вітаміни
Вітамін А (антиксерофтальмічний фактор)
Вітамін А міститься тільки в тваринних продуктах. В рослинах містяться його попередники – провітаміни. Провітамінами вітаміну А є рослинні пігменти каротини (від лат. carota – морква).
Розрізняють 3 різновиди каротинів: альфа, бета і гама.
Обмін вітаміну А
Добова потреба вітаміну А для дорослої людини дорівнює в середньому 1,5-2,0 мг (5-6 тисяч МО), для дітей до 1 року – 0,5 мг.
Для вагітних жінок та матерів, що годують грудьми немовлят, дозу збільшують вдвоє. При інфекційних хворобах, пораненнях, а також при роботах, пов'язаних із напруженням зору, кількість вітаміну А також треба збільшувати.
Найбільше вітаміну А міститься в печінці морських риб, в печінці рогатої худоби, нирках і дещо менше у яєчному жовтку, вершковому маслі. Відповідно каротину багато є в червоному перці, моркві, цибулі, салаті, шпинаті, капусті, помідорах, ягодах обліпихи, горобини, абрикосах. Для всмоктування вітаміну А, як і інших жиророзчинних вітамінів, необхідні жовчні кислоти та жири. Каротини в слизовій кишечника і печінці частково перетворюються у вітамін А. У слизовій кишечника ретинол утворює складні ефіри з жирними кислотами, що мають довгий вуглецевий ланцюг. Ці ефіри транспортуються в складі хіломікронів через лімфатичну систему в кров. У плазмі крові ретинол зв'язується з ретинолтранспортним білком (фракція альфа1-глобулінів) і доставляється до тканин. У сітківці ретинол перетворюється в ретиналь, що входить до складу родопсину і бере участь у зоровому процесі. Відкладається ретинол у печінці, частина його тут окиснюється в ретиналь, далі в ретиноєву кислоту, яка виділяється із жовчю у вигляді глюкуронідів. Усі форми вітаміну А (ретинол, ретиналь, ретиноєва кислота та ефірні форми) беруть участь у біохімічних процесах організму. За рахунок подвійних зв'язків вітамін А має відношення до регуляції окисно-відновних процесів, легко окиснюючись у складі мембран, змінює їх проникність та біосинтез компонентів мембран. Він стимулює бар'єрну функцію та проліферацію шкіри і всіх слизових, загальмовує перетворення циліндричного епітелію в плоский зроговілий; регулює нормальний ріст та диференціацію клітин ембріона і молодого організму, сприяє нормальному розвитку сперматозоїдів та плаценти під час вагітності. Реалізація цих функцій здійснюється за допомогою всіх форм вітаміну А, крім ретиноєвої кислоти. Ретиноєва кислота стимулює ріст кісток та м'яких тканин, що реалізується через активацію експресії генів. В ядрі клітин ретиноєва кислота зв'язується із внутрішньоядерним рецептором (рис. ). Утворений комплекс взаємодіє з хроматином, активує транскрипцію відповідних генів, що призводить до синтезу відповідних білків.
Але цілковито механізми участі вітаміну А в метаболічних процесах, що забезпечують його функції, не досліджені. Найбільш вивчена роль вітаміну А в акті зору. В цьому процесі він бере участь у формі цис-ретиналю – компонента, що входить до складу білка родопсину. Родопсин – хромопротеїн, що складається з білка опсину і простетичної групи – ретиналю. Відіграє роль фоторецептора плазматичних мембран світлочутливих клітин сітківки ока. У людини сітківка ока має 2 види світлочутливих клітин – палички і колбочки. Палички сприймають слабке освітлення (забезпечують сутінковий і нічний зір), колбочки реагують на добре освітлення (забезпечують кольоровий і денний зір). У колбочках замість родопсину є йодопсин, вони відрізняються між собою білковою частиною.
Під впливом світла ретиналь змінюється (із цис- переходить у трансформу). Одночасно поглинутий квант світла призводить до дисоціації родопсину на опсин та транс-ретиналь, що супроводжується деполяризацією мембран та виникненням потенціалу дії. Останній, поширюючись по зоровому нерву, зоровому перехресті, несе інформацію в мозок, у ділянку fissura calcarina, де здійснюється первинний аналіз світловідчуття (вищий аналіз відбувається у корі головного мозку цієї ділянки).
Відновлення родопсину здійснюється в темряві. Тут під впливом алкогольдегідрогенази за участю коферменту НАД транс-ретиналь відновлюється в транс-ретинол. Останній за допомогою ізомерази перетворюється в цис-ретинол, який через посередництво вже згаданої алкогольдегідрогенази окиснюється в цис-ретиналь. Цис-ретиналь, взаємодіючи з опсином, регенерує родопсин, який може далі брати участь у сприйнятті світлового сигналу. Перетворення родопсину на світлі і в темряві у вигляді схеми показані на таблиці. Існує думка, що транс-ретиналь може перетворюватися на світлі в цис-ретиналь під впливом ізомерази. Але значення цього процесу другорядне.
Під час відщеплення ретиналю від родопсину частина його руйнується, у зв'язку з чим для ресинтезу родопсину потрібні нові молекули вітаміну А. Тому при відсутності вітаміну А буде обмежений процес утворення цис-ретиналю та родопсину, що проявлятиметься зниженням здатності бачити ввечері та вночі – розвивається "куряча сліпота" (гемералопія), тобто знижується темнова адаптація, що є ранньою ознакою гіповітамінозу А. При гіповітамінозі порушується структура і функція епітелію – він орогіває, злущується. Це проявляється сухістю шкіри (ксеродермія), ороговінням епідермісу (гіперкератоз). Внаслідок ороговіння циліндричного епітелію та закупорювання слізних каналів настає сухість кон'юнктиви ока (ксерофтальмія), яка під впливом інфекції може швидко призводити до розм'якшення рогівки (кератомаляція):
Одночасно спостерігається ороговіння та злущення епітелію дихальних шляхів, травного та сечовидільного трактів, що нерідко супроводжується закупорюванням видільних проток.
Хронічна нестача вітаміну А в молодому віці проявляється сповільненням росту, частими запаленнями дихальних шляхів. Надмірне потрапляння в організм вітаміну А супроводжується збільшенням проникності або руйнуванням мембран. Тому із лізосом в навколишнє середовище виходять кислі протеази та кисла фосфатаза, із мітохондрій – малатдегідрогеназа. Змін зазнають і ядерні мембрани. Гострий гіпервітаміноз А проявляється важкою інтоксикацією. Як антагоніст вітаміну А призначають тироксин та аскорбінову кислоту.
Вітамін D (кальциферол, антирахітний)
У шкірі під дією ультрафіолетового випромінювання 7‑дегідрохолестерин перетворюється в холекальциферол, з нього у печінці утворюється 25-гідроксихолекальциферол, а в нирках – 1,25-дигідроксихолекальциферол. Таких перетворень зазнає як вітамін D2, так і вітамін D3. На обмін речовин вітамін D діє як синергіст паратгормону та антагоніст гормону щитовидної залози – тиреокальцитоніну. Є навіть думка, що активна форма вітаміну D є складовим ліпідним компонентом, який зв'язується з білковою частиною паратгормону. На рис. показана роль вітаміну D3 в обміні кальцію і фосфору.
Активна форма вітаміну D підтримує постійний рівень кальцію і фосфору в крові. У крові здорових людей вміст Са в середньому складає 2,2-2,7 ммоль/л, фосфору – 1,2-2,2 ммоль/л. Вітамін D стимулює всмоктування кальцію і фосфору в кишечнику за допомогою кишкової Са2+-АТФази та кальційзв'язувального білка.
Нестача вітаміну D в організмі дітей проявляється у вигляді захворювання, яке називається рахітом. В основі цього захворювання лежать зміни обміну фосфору і кальцію, порушення відкладання їх солей у кістковій тканині. Чим молодший вік дитини, тим вища можливість захворювання на рахіт. Це пов'язано з тим, що молодші діти (грудного віку) одержують менше вітаміну D з їжею та меншу дозу ультрафіолетового опромінення. При рахіті загальмовані процеси всмоктування іонів кальцію і фосфатів у кишечнику і реабсорбція їх у нирках. Тому рівень їх у крові знижується (фосфору – на 50 %, кальцію – на 30 % від норми), загальмовується мінералізація кісток, тобто не відбувається відкладання мінеральних речовин на колагенову основу кісток, що ростуть. При рахіті зростає (більш як на порядок) вміст лужної фосфатази крові. Можливо, що це є компенсаторна реакція, спрямована на вирівнювання зниженого вмісту неорганічного фосфату.
Раннім проявом гіповітамінозу D є функціональні розлади центральної нервової системи в дитини, що проявляється переважанням процесів збудження. Трохи пізніше зміни торкаються м'язової системи (знижується тонус м'язів) та кістково-хрящового апарату.
Добова доза вітаміну D для дітей знаходиться в межах 12-25 мкг (500-1000 МО). Для дорослої людини потреба у вітаміні D в десятки разів менша, ніж для дітей. Джерелом вітаміну D є риб'ячий жир, печінка, вершкове масло, жовток яйця.
Вітамін Е (токоферол, вітамін розмноження)
Вітамін Е для людини і вищих тварин є потужним і головним жиророзчинним антиоксидантом. Його дія спрямована на посилення тканинного дихання й утримування на стаціонарному рівні вільнорадикального перекисного окиснення. Опосередковано як кофактор бере участь у транспорті електронів і протонів у дихальному ланцюзі, стимулює синтез убіхінону. Токоферол служить "пасткою" для вільних радикалів – утворює з ними неактивні форми, які обривають вільнорадикальний ланцюг. Цим він захищає від перекисного окиснення поліненасичені жирні кислоти в складі клітинних мембран. Мембранозахисна функція токоферолу тісно пов'язана із біологічною роллю селену, який є кофактором глутатіонпероксидази, що руйнує пероксидрадикали. Токоферол, інактивуючи першим вільні радикали, знижує потребу в глутатіонпероксидазі. Він також підвищує біологічну активність вітаміну А, захищаючи його боковий ненасичений ланцюг від окиснення. Звідси зрозуміло, чому саме гіповітаміноз Е супроводжується патологією мембран у вигляді схильності еритроцитів до перекисного гемолізу, атрофії сім'яників, що призводить до безпліддя, розсмоктування плода під час вагітності, м'язової дистрофії, втрати внутрішньоклітинних компонентів та білків м'язів, некрозу печінки, розм'якшення мозку. Гіпервітамінозу Е в людини не виявлено. Відомо, що при надмірному надходженні вітаміну Е в організмі активується пероксидне окиснення ліпідів, яке приводить до підвищення використання вітаміну Е та зниження його вмісту до нормального рівня.
Вітамін К
(нафтохінони, антигеморагічний,
або вітамін коагуляції)
До вітаміну К відносять дві групи хінонів, в яких бічні радикали – ізопренові ланцюги різної довжини. Існує синтетичний аналог вітаміну К, який не містить бічного ланцюга в 3-му положенні. Його називають вітаміном К3 (2-метил-1,4-нафтохінон). На його основі були одержані інші водорозчинні препарати з вітамінною дією, зокрема вікасол, який являє собою натрієву сіль бісульфітного похідного вітаміну К3.
Вітамін К – антигеморагічний фактор, що має пряме відношення до згортання крові. При нестачі вітаміну К виникають геморагії (крововиливи).