БИОХИМИЧЕСКИЕ ФУНКЦИИ ПЕЧЕНИ. МЕТАБОЛИЗМ
ПОРФИРИНА: ОБМЕН ЖЕЛЧНЫХ ПИГМЕНТОВ, БИОХИМИЯ ЖЕЛТУХ. БИОТРАНСФОРМАЦИЯ
КСЕНОБИОТИКОВ И ЭНДОГЕННЫХ ТОКСИНОВ В
ПЕЧЕНИ: МИКРОСОМАЛЬНОЕ ОКИСЛЕНИЕ, ЦИТОХРОМ Р-450
ПЕЧЕНЬ

Важнейшее
значение печени в обмене
веществ в первую очередь определяется тем, что она является как бы
большой промежуточной станцией между портальным и общим кругом кровообращения.
http://www.youtube.com/watch?v=tat0QYxlCbo&feature=related
В печень
человека более 70% крови поступает через воротную вену, остальная кровь
попадает через печеночную артерию. Кровь воротной вены омывает всасывающую
поверхность кишечника, и в результате большая часть веществ, всасывающихся в
кишечнике, проходит через печень (кроме липидов, транспорт которых
в основном осуществляется через лимфатическую систему). Таким образом, печень
функционирует как первичный регулятор содержания в крови веществ, поступающих в организм с пищей.
Доказательством справедливости данного положения является следующий общий факт:
несмотря на то что всасывание питательных веществ из кишечника в
кровь происходит прерывисто, непостоянно, в связи с чем в портальном круге
кровообращения могут наблюдаться изменения концентрации ряда веществ (глюкоза, аминокислоты и др.), в
общем круге кровообращения изменения в концентрации указанных
соединений незначительны. Все это подтверждает важную роль печени в поддержании
постоянства внутренней среды организма. Печень
выполняет также крайне важную экскреторную функцию, теснейшим образом связанную
с ее детоксикационной функцией. В целом без преувеличения можно констатировать,
что в организме
нет путей обмена
веществ, которые прямо или косвенно не контролировались бы печенью,
в связи с чем многие важнейшие функции печени уже рассматривались в
соответствующих главах учебника. В данной главе будет сделана попытка дать
обобщающие представления о роли печени в обмене веществ целостного организма.
ХИМИЧЕСКИЙ СОСТАВ ПЕЧЕНИ
У взрослого здорового человека масса печени составляет в среднем 1,5 кг. Некоторые исследователи
считают, что эту величину следует рассматривать как нижнюю границу нормы, а
диапазон колебаний от 20 до 60
г на 1
кг массы тела. В табл. представлены некоторые данные о
химическом составе печени в норме. Из данных табл. видно, что более 70% от
массы печени составляет вода.
Однако следует помнить, что масса печени и ее состав подвержены значительным
колебаниям как в норме, так и особенно при патологических состояниях. Например,
при отеках количество воды
может составлять до 80% от массы печени, а при избыточном отложении жира в печени – снизиться
до 55%. Более половины сухого остатка печени приходится на долю белков, причем примерно
90% из них – на глобулины.
Печень богата различными ферментами.
Около 5% от массы печени составляют липиды: нейтральные жиры (триглицериды), фосфолипиды, холестерин и др. При
выраженном ожирении
содержание липидов
может достигать 20% от массы органа, а при жировом перерождении печени
количество липидов
может составлять 50% от сырой массы.
В печени может содержаться 150–200 г гликогена. Как правило,
при тяжелых паренхиматозных поражениях печени количество гликогена в ней
уменьшается. Напротив, при некоторых гликогенозах содержание гликогена достигает 20% и
более от массы печени.
Разнообразен и минеральный состав печени. Количество железа, меди, марганца, никеля и некоторых других
элементов превышает их содержание в других органах и тканях.
ПЕЧЕНИ В
УГЛЕВОДНОМ ОБМЕНЕ
Основная роль печени в углеводном обмене заключается в обеспечении
постоянства концентрации
глюкозы в крови. Это
достигается регуляцией между синтезом и распадом гликогена, депонируемого в
печени.
В печени синтез гликогена
и его регуляция в основном аналогичны тем процессам, которые протекают в других
органах и тканях,
в частности в мышечной ткани.
Синтез гликогена
из глюкозы
обеспечивает в норме временный резерв углеводов, необходимый для
поддержания концентрации
глюкозы в крови в тех
случаях, если ее содержание значительно уменьшается (например, у человека это
происходит при недостаточном поступлении углеводов с пищей или в
период ночного «голодания»).
Синтез и распад гликогена
http://www.youtube.com/watch?v=p-lFJVOkFwk
Необходимо подчеркнуть важную роль фермента глюкокиназы в
процессе утилизации глюкозы
печенью. Глюкокиназа, подобно гексокиназе, катализирует фосфорилирование глюкозы с образованием
глюкозо-6-фосфата, при этом активность
глюкокиназы в печени почти в 10 раз превышает активность гексокиназы.
http://www.youtube.com/watch?v=O5eMW4b29rg&feature=related
Важное различие между этими двумя ферментами заключается в
том, что глюкокиназа в противоположность гексокиназе имеет высокое
значение КМ для глюкозы
и не ингибируется глюкозо-6-фосфатом.
Синтез гликогена
Распад гликогена
После приема
пищи содержание глюкозы
в воротной вене резко возрастает: в тех же пределах увеличивается и ее
внутрипеченочная концентрация
. Повышение концентрации
глюкозы в печени вызывает
существенное увеличение активности
глюкокиназы и автоматически увеличивает поглощение глюкозы печенью
(образовавшийся глюкозо-6-фосфат либо затрачивается на синтез гликогена, либо
расщепляется).
|
Механизм действия фосфорилазы гликогена
|
Особенности метаболизма гликогена в печени и мышцах
http://www.youtube.com/watch?v=oBL0OC3IavI
Считают, что
основная роль печени – расщепление глюкозы – сводится прежде
всего к запасанию метаболитов-предшественников, необходимых для биосинтеза жирных кислот и
глицерина, и в меньшей
степени к окислению
ее до СО2 и Н2О. Синтезированные в печени триглицериды в
норме выделяются в кровь в составе липопротеинов и
транспортируются в жировую ткань
для более «постоянного» хранения.
В реакциях
пентозофосфатного пути в печени образуется НАДФН, используемый для
восстановительных реакций в процессах синтеза жирных кислот, холестерина и других стероидов. Кроме того, при
этом образуются пентозофосфаты, необходимые для синтеза нуклеиновых кислот.
Пентозофосфатный путь превращения глюкозы
Наряду с
утилизацией глюкозы
в печени происходит и ее образование. Непосредственным источником глюкозы в печени служит гликоген. Распад гликогена в печени
происходит в основном фосфоролитическим путем. В регуляции скорости
гликогенолиза в печени большое значение имеет система циклических нуклеотидов. Кроме того, глюкоза в печени
образуется также в процессе глюконеогенеза.
Глюконеогенез. Ферменты: 1- пируваткарбоксилаза, 2-
фосфоенолпируваткарбоксикиназа, 3- фосфатаза фру-1,6-дифосфата, 4-
глюкозо-6-фосфатаза.
Основными субстратами глюконеогенеза служат
лактат, глицерин
и аминокислоты. Принято
считать, что почти все аминокислоты,
за исключением лейцина,
могут пополнять пул предшественников
глюконеогенеза.
При оценке
углеводной функции печени необходимо иметь в виду, что соотношение между
процессами утилизации и образования глюкозы регулируется
прежде всего нейрогуморальным путем при участии желез внутренней секреции.
Центральную
роль в превращениях глюкозы
и саморегуляции углеводного обмена в печени играет глюкозо-6-фосфат. Он резко
тормозит фосфоролитическое расщепление гликогена, активирует
ферментативный перенос глюкозы
с уридиндифосфоглюкозы на молекулу
синтезирующегося гликогена,
является субстратом
для дальнейших гликолитических превращений, а также окисления глюкозы, в том числе по
пентозофосфатному пути. Наконец, расщепление глюкозо-6-фосфата фосфатазой обеспечивает
поступление в кровь свободной глюкозы, доставляемой
током крови во все органы и ткани
(рис. ).
Рис. Участие глюкозо-6-фосфата в метаболизме углеводов.
Рис. Гормональная регуляция системы фруктозо-2,6-бисфосфата
(Ф-2,6-Р2) в печени при участии цАМФ-зависимых протеинкиназ.
http://www.youtube.com/watch?v=oBL0OC3IavI
Как
отмечалось, наиболее мощным аллостерическим активатором фосфофруктокиназы-1 и ингибитором
фруктозо-1,6-бисфосфатазы печени является фруктозо-2,6-бисфосфат (Ф-2,6-Р2).
Повышение в гепатоцитах уровня Ф-2,6-Р2 способствует усилению гликолиза и уменьшению
скорости глюконеогенеза.
Ф-2,6-Р2 снижает ингибирующее действие АТФ на
фосфо-фруктокиназу-1 и увеличивает сродство этого фермента к
фруктозо-6-фосфату. При ингибировании фруктозо-1,6-бисфосфатазы Ф-2,6-Р2
возрастает значение КМ для фруктозо-1,6-бисфосфата. Содержание
Ф-2,6-Р2 в печени, сердце, скелетной мускулатуре и других тканях контролируется
бифункциональным ферментом,
который осуществляет синтез Ф-2,6-Р2 из фруктозо-6-фосфата и АТФ и гидролиз его до
фруктозо-6-фосфата и Pi, т.е. фермент одновременно
обладает и киназной, и бисфосфатазной активностью.
Бифункциональный фермент
(фосфофруктокиназа-2/фруктозо-2,6-бисфосфатаза), выделенный из печени крысы,
состоит из двух идентичных субъединиц с мол. массой 55000, каждая из которых
имеет два различных каталитических центра. Киназный домен при этом расположен
на N-конце, а бисфосфатазный – на С-конце каждой из полипептидных цепей.
Известно также, что бифункциональный фермент печени является
прекрасным субстратом
для цАМФ-зависимой протеинкиназы
А. Под действием протеинкиназы А происходит фосфорилирование остатков серина в каждой из
субъединиц бифункционального фермента, что приводит к
снижению его киназной и повышению бисфосфатазной активности. Заметим, что в
регуляции активности
бифункционального фермента
существенная роль принадлежит гормонам, в частности глюкагону.
При многих
патологических состояниях, в частности при сахарном диабете, отмечаются
существенные изменения в функционировании и регуляции системы Ф-2,6-Р2.
Установлено, что при экспериментальном (стептозотоциновом) диабете у крыс на
фоне резкого увеличения уровня глюкозы в крови и моче в
гепатоцитах содержание Ф-2,6-Р2 снижено. Следовательно, снижается
скорость гликолиза
и усиливается глюконео-генез. Данный факт имеет свое объяснение. Возникающие у
крыс при диабете нарушения гормонального фона: увеличение концентрации глю-кагона и
уменьшение содержания инсулина
– обусловливают повышение концентрации
цАМФ в ткани
печени, усиление цАМФ-зависимого фосфорилирования
бифункционального фермента,
что в свою очередь приводит к снижению его киназной и повышению бисфосфатазной активности. Таков может
быть механизм снижения уровня Ф-2,6-Р2 в гепатоцитах при
экспериментальном диабете. По-видимому, существуют и другие механизмы, ведущие
к снижению уровня Ф-2,6-Р2 в гепатоцитах при стрептозото-циновом
диабете. Показано, что при экспериментальном диабете в ткани печени имеет место
снижение активности
глюкокиназы (возможно, и снижение количества данного фермента). Это приводит к
падению скорости фосфорилирования
глюкозы, а затем к
снижению содержания фруктозо-6-фосфата – субстрата
бифункционального фермента.
Наконец, в последние годы было показано, что при стрептозотоциновом диабете
уменьшается количество мРНК
бифункционального фермента
в гепатоцитах и как следствие – снижается уровень Ф-2,6-Р2 в ткани печени, усиливается
глюко-неогенез. Все это еще раз подтверждает положение, что Ф-2,6-Р2,
являясь важным компонентом в цепи передачи гормонального сигнала, выступает в
роли третичного посредника при действии гормонов, прежде всего на
процессы гликолиза
и глюконеогенеза.
Рассматривая промежуточный
обмен углеводов
в печени, необходимо также остановиться на превращениях фруктозы и галактозы. Поступающая в
печень фруктоза
может фосфорилироваться в положении 6 до фруктозо-6-фосфата под действием гексокиназы, обладающей
относительной специфичностью
и катализирующей фосфорилирование,
кроме глюкозы
и фруктозы,
еще и маннозы.
Однако в печени существует и другой путь: фруктоза способна
фосфорилироваться при участии более специфического фермента – фруктокиназы. В
результате образуется фруктозо-1-фосфат. Эта реакция не блокируется глюкозой. Далее
фруктозо-1-фосфат под действием альдолазы расщепляется на
две триозы: диоксиацетонфосфат и глицераль-дегид. Под влиянием соответствующей
киназы (триокиназы) и при участии АТФ глицеральдегид
подвергается фосфорилированию
до глицеральдегид-3-фосфата. Последний (в него легко переходит и
диоксиацетонфосфат) подвергается обычным превращениям, в том числе с
образованием в качестве промежуточного продукта пировиноградной кислоты.
Связь пентозофосфатного пути превращения глюкозы с гликолизом и
глюконеогенезом
Следует
отметить, что при генетически обусловленной нетолерантности к фруктозе или недостаточной
активности
фруктозо-1,6-бисфосфатазы наблюдается индуцируемая фруктозой гипогликемия,
возникающая вопреки наличию больших запасов гликогена. Вероятно,
фруктозо-1-фосфат и фруктозо-1,6-бисфосфат ингибируют фосфорилазу печени по
аллосте-рическому механизму.
Известно также, что метаболизм
фруктозы по
гликолитическому пути в печени происходит гораздо быстрее, чем метаболизм глюкозы. Для метаболизма глюкозы характерна стадия,
катализируемая фосфофрукто-киназой-1. Как известно, на этой стадии
осуществляется метаболический контроль скорости катаболизма глюкозы. Фруктоза минует эту стадию,
что позволяет ей интенсифицировать в печени процессы метаболизма, ведущие к
синтезу жирных кислот, их эстерификацию и секрецию липопротеинов очень низкой
плотности; в результате может увеличиваться концентрация триглицеридов
в плазме крови.
Галактоза
в печени сначала фосфорилируется при участии АТФ и фермента галактокиназы с
образованием галактозо-1-фосфата. Для га-лактокиназы печени плода и ребенка характерны
значения КМ и Vмaкс, примерно в 5 раз превосходящие
таковые у ферментов
взрослого человека. Большая часть галактозо-1-фосфата в печени превращается в
ходе реакции, катализируемой гексозо-1-фосфат-уридилилтрансферазой:
УДФ-глюкоза + Галактозо-1-фосфат –> УДФ-галактоза + Глюкозо-1-фосфат.
Это уникальная трансферазная реакция возвращения галактозы в основное русло
углеводного метаболизма.
Наследственная утрата
гексозо-1-фосфат-уридилилтрансферазы приводит к галактоземии – заболеванию, для
которого характерны умственная отсталость и катаракта хрусталика. В этом случае
печень новорожденных теряет способность метаболизи-ровать D-галактозу, входящую
в состав лактозы
молока.
РЕГУЛЯЦИЯ ОБМЕНА ОСНОВНЫХ ЭНЕРГОНОСИТЕЛЕЙ ПРИ НОРМАЛЬНОМ
РИТМЕ ПИТАНИЯ
1. Энергетическая ценность основных пищевых веществ выражается в
килокалориях и составляет: для углеводов - 4 ккал/г, для жиров - 9 ккал/г, для
белков - 4 ккал/г. Взрослому здоровому человеку в сутки требуется 2000- 3000
ккал (8000-12 000 кДж) энергии.
При обычном ритме питания промежутки между приемами пищи
составляют 4-5 часов с 8-12-часовым ночным перерывом. В течение пищеварения и абсорбтивного
периода (2-4 часа) основные энергоносители, используемые тканями (глюкоза,
жирные кислоты, аминокислоты), могут поступать в кровь непосредственно из
пищеварительного тракта. В постабсорбтивном периоде (промежуток времени
после завершения пищеварения до следующего приема пищи) и при голодании
энергетические субстраты образуются в процессе катаболизма депонированных энергоносителей.
Основную роль в регуляции этих процессов играют инсулин и глюкагон. Антагонистами
инсулина являются также адреналин, кортизол, иодтиронины и соматотропин (так называемые контринсулярные
гормоны).
Инсулин и контринсулярные гормоны обеспечивают баланс между
потребностями и возможностями организма в получении энергии, необходимой для
нормального функционирования и роста. Этот баланс определяется как энергетический
гомеостаз. При нормальном ритме питания концентрация глюкозы в крови
поддерживается на уровне 65-110 мг/дл (3,58-6,05 ммоль/л) благодаря влиянию
двух основных гормонов - инсулина и глюкагона. Инсулин и глюкагон - главные
регуляторы метаболизма при смене состояний пищеварения, постабсорбтивного
периода и голодания. На периоды пищеварения приходится 10-15 час в сутки, а
расход энергии происходит в течение 24 часов. Поэтому часть энергоносителей во
время пищеварения запасается для использования в постабсорбтивном периоде.
Печень, жировая ткань и мышцы - главные органы,
обеспечивающие изменения метаболизма в соответствии с ритмом питания. Режим
запасания включается после приема пищи и сменяется режимом мобилизации запасов
после завершения абсорбтивного периода.
2. Изменения метаболизма основных энергоносителей в
абсорбтивном периоде обусловлены, в основном, высоким инсулин-глюкагоновым индексом (рис.).
Рис. Пути использования
основных энергоносителей в абсорбтивном периоде: 1 - биосинтез гликогена в
печени; 2 - гликолиз; 3 - биосинтез ТАГ в печени; 4 - биосинтез ТАГ в жировой
ткани; 5 - биосинтез гликогена в мышцах; 6 - биосинтез белков в разных тканях,
в том числе в печени; ЖК - жирные кислоты
Изменения метаболизма в печени. В печени увеличивается потребление
глюкозы, что является следствием ускорения метаболических путей, в которых
глюкоза превращается в депонируемые формы энергоносителей: гликоген и жиры.
При повышении концентрации глюкозы в гепатоцитах происходит
активация глюкокиназы, превращающей глюкозу в глюкозо-6-фосфат. Кроме этого,
инсулин индуцирует синтез мРНК глюкокиназы. В результате повышается
концентрация глюкозо-6-фосфата в гепатоцитах, что обусловливает ускорение синтеза
гликогена. Этому также способствует одновременная инактивация
гликогенфосфорилазы и активация гликогенсинтазы. Под влиянием инсулина в
гепатоцитах ускоряется гликолиз в результате повышения активности и
количества ключевых ферментов: глюкокиназы, фосфофруктокиназы и пируваткиназы.
В то же время происходит торможение глюконеогенеза в результате инактивации
фруктозо-1,6-бисфосфатазы и репрессии инсулином синтеза
фосфоенолпируваткарбоксикиназы - ключевых ферментов глюконеогенеза.
Повышение концентрации глюкозо-6-фосфата в гепатоцитах в
абсорбтивном периоде сочетается с активным использованием NADPH для синтеза
жирных кислот, что способствует стимуляции пентозофосфатного пути.
Ускорение синтеза жирных кислот обеспечивается доступностью
субстратов (ацетил-КоА и NADPH), образующихся при метаболизме глюкозы, а также
активацией и индукцией ключевых ферментов синтеза жирных кислот инсулином.
Аминокислоты, поступающие в печень из пищеварительного
тракта, используются для синтеза белков и других азотсодержащих соединений, а
их излишек либо поступает в кровь и транспортируется в другие ткани, либо
дезаминируется с последующим включением безазотистых остатков в общий путь
катаболизма.
Изменения метаболизма в адипоцитах. Основная функция жировой ткани -
запасание энергоносителей в форме триацилглицеролов. Под влиянием
инсулина ускоряется транспорт глюкозы в адипоциты. Повышение
внутриклеточной концентрации глюкозы и активация ключевых ферментов гликолиза
обеспечивают образование ацетил-КоА и глицерол-3-фосфата, необходимых для
синтеза ТАГ. Стимуляция пентозофосфатного пути обеспечивает образование NADPH,
необходимого для синтеза жирных кислот. Однако биосинтез жирных кислот de novo
в жировой ткани человека протекает с высокой скоростью только после
предшествующего голодания. При нормальном ритме питания для синтеза ТАГ
используются в основном жирные кислоты, поступающие из хиломикронов и ЛПОНП под
действием ЛП-липазы.
Так как гормончувствительная ТАГ-липаза в абсорбтивном
состоянии находится в дефосфорилированной, неактивной форме, процесс липолиза
тормозится.
Изменение метаболизма в мышцах. Под влиянием инсулина ускоряется транспорт
глюкозы в клетки мышечной ткани. Глюкоза фосфорилируется и окисляется для
обеспечения клеток энергией, а также используется для синтеза гликогена. Жирные
кислоты, поступающие из хиломикронов и ЛПОНП, в этот период играют
незначительную роль в энергетическом обмене мышц. Поток аминокислот в мышцы и
биосинтез белков также возрастает под влиянием инсулина, особенно после приема
белковой пищи и в период мышечной работы.
3. Изменения метаболизма основных энергоносителей при смене
абсорбтивного состояния на постабсорбтивное. В постабсорбтивном периоде при
снижении инсулин-глюкагонового индекса изменения метаболизма направлены главным
образом на поддержание концентрации в крови глюкозы, которая служит главным
энергетическим субстратом для мозга и единственным источником энергии для
эритроцитов. Основные изменения метаболизма в этот период происходят в печени и
жировой ткани и направлены на пополнение глюкозы за счет внутренних резервов и
на использование других энергетических субстратов (жиров и аминокислот) (рис.).
Рис. Пути использования
основных энергоносителей при смене абсорбтивного состояния на постабсорбтивное: I - снижение
инсулин-глюкагонового индекса; 2 - распад гликогена; 3, 4 - транспорт глюкозы в
мозг и эритроциты; 5 - катаболизм жиров; 6 - транспорт жиров в печень и мышцы;
7 - синтез кетоновых тел в печени; 8 - транспорт кетоновых тел в мышцы; 9 -
глюконеогенез из аминокислот; 10 - синтез и выведение мочевины; II - транспорт лактата в
печень и включение в глюконеогенез; 12 - глюконеогенез из глицерола; КТ
-кетоновые тела; ЖК - жирные кислоты
Изменения метаболизма в печени. Под влиянием глюкагона
ускоряется мобилизация гликогена. Запасы гликогена в печени истощаются в
течение 18-24-часового голодания. Главным источником глюкозы по мере исчерпания
запасов гликогена становится глюконеогенез, который начинает ускоряться
через 4-6 часов после последнего приема пищи. Субстратами для синтеза глюкозы
служат лактат, глицерол и аминокислоты. Скорость синтеза жирных
кислот снижается вследствие фосфорилирования и инактивации
ацетил-КоА-карбоксилазы при фосфорилировании, а скорость β-окисления
возрастает. Вместе с тем увеличивается снабжение печени жирными кислотами,
которые транспортируются из жировых депо в результате ускорения липолиза.
Ацетил-КоА, образующийся при окислении жирных кислот, используется в печени для
синтеза кетоновых тел.
В жировой ткани снижается скорость синтеза ТАГ
и стимулируется липолиз. Стимуляция липолиза является результатом активации
гормончувствительной ТАГ-липазы адипоцитов под влиянием глюкагона. Жирные
кислоты становятся важными источниками энергиии в печени, мышцах и жировой
ткани.
Таким образом, в постабсорбтивном
периоде концентрация глюкозы в крови поддерживается на уровне 60-100 мг/дл
(3,5-5,5 ммоль/л), а уровень жирных кислот и кетоновых тел возрастает.
РОЛЬ ПЕЧЕНИ В ЛИПИДНОМ ОБМЕНЕ
Рис. .
Участие печени в жировом обмене
Рис. Основные
этапы метаболизма липидов в гепатоците
Ферментные
системы печени способны катализировать все реакции или значительное большинство
реакций метаболизма
липидов. Совокупность этих
реакций лежит в основе таких процессов, как синтез высших жирных кислот,
триглицеридов, фосфолипидов,
холестерина и его эфиров,
а также липолиз триглицеридов, окисление жирных кислот,
образование ацетоновых (кетоновых) тел и т.д. Напомним, что ферментативные
реакции синтеза триглицеридов в печени и жировой ткани сходны. Так,
КоА-производные жирной кислоты с длинной цепью взаимодействуют с
глицерол-3-фосфатом с образованием фосфатидной кислоты, которая затем
гидролизуется до диглицерида. Путем присоединения к последнему еще одной молекулы КоА-производного жирной
кислоты образуется триглицерид. Синтезированные в печени три-глицериды либо
остаются в печени, либо секретируются в кровь в форме липопротеинов. Секреция происходит с
известной задержкой (у человека 1–3 ч). Задержка секреции, вероятно,
соответствует времени, необходимому для образования липопротеинов. Основным
местом образования плазменных пре-β-липопротеинов (липопротеины очень
низкой плотности – ЛПОНП) и α-липопротеинов (липопротеины высокой
плотности – ЛПВП) является печень.
ХС - холестерин
ТГ - триглицериды
ЖК - жирные кислоты
ХМ - хиломикроны
ЛПЛ - липопротеинлипаза
ТГЛ - триглицеридлипаза.
http://www.youtube.com/watch?v=97uiV4RiSAY
Рассмотрим образование ЛПОНП. Согласно данным литературы, основной белок апопротеин В-100
(апо Б-100) липопротеинов
синтезируется в рибосомах
шероховатого эндоплазматического ретикулума гепатоцитов. В гладком
эндоплазматическом ретикулуме, где синтезируются и липидные компоненты,
происходит сборка ЛПОНП. Одним из основных стимулов образования ЛПОНП является
повышение концентрации
неэстерифици-рованных жирных кислот (НЭЖК). Последние либо поступают в печень с
током крови, будучи связанными с альбумином, либо
синтезируются непосредственно в печени. НЭЖК служат главным источником
образования триглицеридов (ТГ). Информация о наличии НЭЖК и ТГ передается на
мембранно-связанные рибосомы
шероховатого эндоплазматического ретикулума, что в свою очередь является
сигналом для синтеза белка
(апо В-100). Синтезированный белок
внедряется в мембрану
шероховатого ретикулума, и после взаимодействия с фосфолипидным бислоем от мембраны отделяется
участок, состоящий из фосфолипидов
(ФЛ) и белка,
который и является предшественником
ЛП-частицы. Далее белокфосфо-липидный комплекс поступает в гладкий
эндоплазматический ретикулум, где взаимодействует с ТГ и эстерифицированным холестерином (ЭХС), в
результате чего после соответствующих структурных перестроек формируются
насцентные, т.е. незавершенные, частицы (н-ЛПОНП). Последние поступают через
тубулярную сеть аппарата Гольджи в секреторные везикулы и в их составе
доставляются к поверхности клетки,
после чего очень низкой плотности (ЛПОНП) в печеночной клетке (по А.Н. Климову и
Н.Г. Никульчевой) путем экзоцитоза
выделяются в перисинусоидные пространства (пространства Диссе). Из последнего
н-ЛПОНП поступают в просвет кровяного синусоида, где происходят перенос
апопротеинов С из ЛПВП на н-ЛПОНП и достраивание последних (рис. 16.3).
Установлено, что время синтеза апо В-100, образования липид-белковых комплексов
и секреции
готовых частиц ЛПОНП составляет 40 мин.
Рис. Образование липопротеинов
1 - ядро; 2 - шероховатый эндоплазматический ретикулум; 3 - гладкий
эндоплазматический ре-тикулум, синтезированные в нем липиды и образовавшиеся
н-ЛПОНП; 4 - аппарат Гольджи; 5 -секреторная везикула с частицей н-ЛПОНП; 6
-частица с н-ЛПОНП в пространстве Диссе; 7 -перенос апопротеинов С с ЛПВП на
н-ЛПОНП; 8 - частица нативных ЛПОНП.
http://www.youtube.com/watch?v=x-4ZQaiZry8
У человека основная масса β-липопротеинов (липопротеины низкой
плотности – ЛПНП) образуется в плазме крови из ЛПОНП при
действии липопротеинлипазы. В ходе этого процесса образуются сначала
промежуточные короткоживущие липопротеины (Пр.ЛП), а
затем формируются частицы, обедненные триглицеридами и обогащенные холестерином, т.е. ЛПНП.
При высоком содержании жирных кислот в плазме их поглощение
печенью возрастает, усиливается синтез триглицеридов, а также окисление жирных кислот, что
может привести к повышенному образованию кетоновых тел.
Следует подчеркнуть, что кетоновые тела образуются в печени в ходе так
называемого β-гидрокси-β-метилглутарил-КоА пути. Однако существует
мнение, что ацетоацетил-КоА, являющийся исходным соединением при кетогенезе,
может образоваться как непосредственно в ходе β-окисле-ния жирных кислот,
так и в результате конденсации
ацетил-КоА [Марри Р. и др., 1993]. Из печени кетоновые тела током крови доставляются
в ткани и органы (мышцы,
почки, мозг и др.), где они быстро окисляются при участии соответствующих ферментов, т.е. по сравнению
с другими тканями
печень является исключением.
В печени происходит интенсивный распад фосфолипидов, а также их
синтез. Помимо глицерина
и жирных кислот, которые входят в состав нейтральных жиров, для синтеза фосфолипидов необходимы неорганические фосфаты и
азотистые соединения, в частности холин, для синтеза
фосфатидхолина. Неорганические
фосфаты в печени имеются в достаточном количестве. При недостаточном
образовании или недостаточном поступлении в печень холина синтез фосфолипидов из
компонентов нейтрального жира
становится либо невозможным, либо резко снижается и нейтральный жир откладывается в
печени. В этом случае говорят о жировой инфильтрации печени, которая может
затем перейти в ее жировую дистрофию. Иными словами, синтез фосфолипидов лимитируется
количеством азотистых оснований, т.е. для синтеза фосфоглицеридов необходим
либо холин,
либо соединения, которые могут являться донорами метильных групп и
участвовать в образовании холина
(например, метионин). Такие соединения получили название липотропных веществ. Отсюда становится
ясным, почему при жировой инфильтрации печени весьма полезен творог, содержащий
белок казеин, в составе которого
имеется большое количество остатков аминокислоты метионина.
Рассмотрим роль печени в обмене стероидов, в частности холестерина. Часть холестерина поступает в организм с пищей, но
значительно большее количество его синтезируется в печени из ацетил-КоА. Биосинтез холестерина в печени
подавляется экзогенным холестерином,
т.е. получаемым с пищей.
Таким образом, биосинтез
холестерина в печени
регулируется по принципу отрицательной обратной связи. Чем больше холестерина поступает с
пищей, тем меньше его синтезируется в печени, и наоборот. Принято считать, что
действие экзогенного холестерина
на биосинтез его в печени
связано с торможением β-гидрокси-β-метилглутарил-КоА-редуктазной
реакции:
Часть синтезированного в печени холестерина выделяется из организма вместе с желчью,
другая часть превращается в желчные
кислоты и используется в других органах для синтеза стероидных гормонов и иных
соединений.
В печени холестерин
может взаимодействовать с жирными кислотами (в виде ацил-КоА) с образованием
эфиров холестерина.
Синтезированные в печени эфиры холестерина поступают в
кровь, в которой содержится также определенное количество свободного холестерина.
РОЛЬ ПЕЧЕНИ В
ОБМЕНЕ БЕЛКОВ
Печень играет центральную роль в обмене белков. Она выполняет
следующие основные функции: синтез специфических белков плазмы; образование мочевины и мочевой кислоты; синтез холина и креатина; трансаминирование и дезаминирование аминокислот, что весьма
важно для взаимных превращений аминокислот, а также для
процесса глюконеогенеза
и образования кетоновых тел. Все альбумины плазмы, 75–90%
α-глобу-линов и 50% β-глобулинов синтезируются гепатоцитами. Лишь
γ-гло-булины продуцируются не гепатоцитами, а системой макрофагов, к
которой относятся звездчатые ретикулоэндотелиоциты (клетки Купфера). В основном
γ-глобулины образуются в печени. Печень является единственным органом, где
синтезируются такие важные для организма белки, как протромбин, фибриноген, проконвертин и
проакцелерин.
При заболеваниях печени определение фракционного состава белков плазмы (или сыворотки)
крови нередко представляет интерес как в диагностическом, так и в
прогностическом плане. Известно, что патологический процесс в гепатоцитах резко
снижает их синтетические возможности. В результате содержание альбумина в плазме крови резко падает,
что может привести к снижению онкотического давления плазмы крови, развитию
отеков, а затем асцита. Отмечено, что при циррозах печени, протекающих с
явлениями асцита, содержание альбуминов
в сыворотке крови
на 20% ниже, чем при циррозах без асцита.
http://www.youtube.com/watch?v=nXRWkorYFXc
Нарушение синтеза ряда белковых факторов системы свертывания крови
при тяжелых заболеваниях печени может привести
к геморрагическим явлениям.
При
поражениях печени нарушается также процесс дезаминирования
аминокислот,
что способствует увеличению их концентрации
в крови и моче. Так, если в норме содержание азота
аминокислот
в сыворотке
крови составляет примерно 2,9–4,3 ммоль/л, то при тяжелых заболеваниях
печени (атрофические процессы) эта величина возрастает до 21 ммоль/л, что
приводит к аминоацидурии. Например, при острой атрофии печени количество тирозина
в суточном количестве мочи может достигать 2 г (при норме 0,02–0,05 г/сут).
В
организме
образование мочевины
в основном происходит в печени. Синтез мочевины
связан с затратой довольно значительного количества энергии (на образование 1 молекулы
мочевины
расходуется 3 молекулы
АТФ). При заболевании печени, когда количество АТФ
в гепатоцитах уменьшено, синтез мочевины
нарушается. Показательно в этих случаях определение в сыворотке отношения азота
мочевины
к аминоазоту. В норме это отношение равно 2:1, а при тяжелом поражении печени
составляет 1:1.
Большая
часть мочевой
кислоты также образуется в печени, где много фермента
ксантиноксидазы, при участии которого оксипурины (гипо-ксантин и ксантин)
превращаются в мочевую
кислоту. Нельзя забывать о роли печени и в синтезе креатина. Имеются два
источника креатина в организме.
Существует экзогенный креатин, т.е. креатин пищевых продуктов (мясо, печень и
др.), и эндогенный креатин, синтезирующийся в тканях.
Синтез креатина происходит в основном в печени, откуда он с током крови
поступает в мышечную ткань.
Здесь креатин, фосфори-лируясь, превращается в креатинфосфат, а из последнего
образуется креатин.
Чужеродные
вещества
(ксенобиотики) в печени нередко превращаются в менее токсичные и даже
индифферентные вещества.
По-видимому, только в этом смысле можно говорить об «обезвреживании» их в
печени. Происходит это путем окисления,
восстановления,
метилирования,
ацетилирования
и конъюгации с теми или иными веществами.
Необходимо отметить, что в печени окисление,
восстановление
и гидролиз
чужеродных соединений осуществляют в основном микросомальные ферменты.
Наряду с микро-сомальным в печени существует также пероксисомальное окисление.
Пероксисомы – микротельца, обнаруженные в гепатоцитах; их можно рассматривать
как специализированные окислительные органеллы. Эти микротельца содержат
оксидазу мочевой
кислоты, лактатоксидазу, окси-дазу D-аминокислот, а также каталазу.
Последняя катализирует расщепление перекиси водорода,
которая образуется при действии указанных оксидаз; отсюда и название этих
микротелец – пероксисомы. Пероксисо-мальное окисление,
так же как и микросомальное, не сопровождается образованием макроэргических
связей.
В
печени широко представлены также «защитные» синтезы, например синтез мочевины,
в результате которого обезвреживается весьма токсичный аммиак.
В результате гнилостных процессов, протекающих в кишечнике, из тирозина
образуются фенол
и крезол,
а из триптофона – скатол
и индол.
Эти вещества
всасываются и с током крови поступают в печень, где обезвреживаются путем
образования парных соединений с серной или глюкуроновой
кислотой.
Обезвреживание
фенола,
крезола,
скатола
и индола
в печени происходит в результате взаимодействия этих соединений не со свободными
серной и глюкуроновой
кислотами, а с их так называемыми активными формами: ФАФС и УДФГК.
Глюкуроновая
кислота участвует не только в обезвреживании продуктов гниения белковых веществ,
образовавшихся в кишечнике, но и в связывании ряда других токсичных соединений,
образующихся в процессе обмена в тканях.
В частности, свободный, или непрямой, билирубин,
обладающий значительной токсичностью,
в печени взаимодействует с глюкуроновой
кислотой, образуя моно- и диглюкурониды билирубина.
Нормальным метаболитом
является и гиппуровая кислота, образующаяся в печени из бензойной
кислоты и глицина.
Синтез
гиппуровой кислоты у человека протекает преимущественно в печени. Поэтому в
клинической практике довольно часто для выяснения антитоксической функции
печени применяют пробу Квика–Пытеля (при нормальной функциональной способности
почек): после нагрузки бензо-атом натрия
в моче определяют количество образовавшейся гиппуровой кислоты. При
паренхиматозных поражениях печени синтез гиппуровой кислоты снижен.
В
печени широко представлены процессы метилирования.
Так, перед выделением с мочой амид никотиновой
кислоты (витамин РР) метилируется в печени; в результате образуется
N-метилникотинамид. Наряду с метилированием
интенсивно протекают и процессы ацетилирования
. В частности, в печени ацетилированию
подвергаются различные сульфаниламидные
препараты.
Примером
обезвреживания токсичных продуктов в печени путем восстановления
является превращение нитробензола
в парааминофенол. Многие ароматические углеводы
обезвреживаются путем окисления
с образованием соответствующих карбоновых
кислот.
Печень
принимает активное участие в инактивации различных гормонов.
С током крови гормоны
попадают в печень, при этом активность
их в большинстве случаев резко снижается или полностью утрачивается. Так, стероидные
гормоны, подвергаясь микросомальному окислению,
инакти-вируются, превращаясь затем в соответствующие глюкурониды и сульфаты.
Под влиянием аминооксидаз в печени происходит окисление
ка-техоламинов и т.д.
Из
приведенных примеров видно, что печень способна инактивировать ряд
сильнодействующих физиологических и чужеродных (в том числе токсичных) веществ.
Рассмотрим
только гемохромогенные пигменты,
которые образуются в организме
при распаде гемоглобина
(в значительно меньшей степени при распаде миоглобина,
цитохромов
и др.). Распад гемоглобина
протекает в клетках
макрофагов, в частности в звездчатых ретикулоэндотелиоцитах, а также в
гистиоцитах соединительной ткани
любого органа.
Начальным
этапом распада гемоглобина
является разрыв одного метинового мостика с образованием вердоглобина. В
дальнейшем от молекулы
вердоглобина отщепляются атом
железа
и белок
глобин.
В результате образуется биливердин, который представляет собой цепочку из
четырех пиррольных колец, связанных метановыми мостиками. Затем биливердин,
восстанавливаясь, превращается в билирубин
– пигмент,
выделяемый с желчью и поэтому называемый желчным
пигментом. Образовавшийся билирубин
называется непрямым (неконъю-гированным) билирубином.
Он нерастворим в воде,
дает непрямую реакцию с диазореактивом, т.е. реакция протекает только после
предварительной обработки спиртом.
В
печени билирубин
соединяется (конъюгирует) с глюкуроновой
кислотой. Эта реакция катализируется ферментом
УДФ-глюкуронилтранс-феразой, при этом глюкуроновая
кислота вступает в реакцию в активной форме, т.е. в виде УДФГК.
Образующийся глюкуронид билирубина
получил название прямого билирубина
(конъюгированный билирубин). Он растворим в воде
и дает прямую реакцию с диазореактивом. Большая часть билирубина
соединяется с двумя молекулами
глюкуроновой
кислоты, образуя диглюкуронид билирубина:
Рис.
Нормальный обмен уробилиногеновых тел (схема).
Образовавшийся
в печени прямой билирубин
вместе с очень небольшой частью непрямого билирубина
выводится с желчью в тонкую кишку. Здесь от прямого билирубина
отщепляется глюкуроновая
кислота и происходит его восстановление
с последовательным образованием мезобилирубина и мезобилиногена
(уробилиногена). Принято считать, что около 10% билирубина
восстанавливается до мезобилиногена на пути в тонкую кишку, т.е. во
внепеченочных желчных путях и в желчном пузыре. Из тонкой кишки часть
образовавшегося мезобилиногена (уробилиногена) резорбируется через кишечную
стенку, попадает в воротную вену и током крови переносится в печень, где
расщепляется полностью до ди- и трипирролов. Таким образом, в норме в общий
круг кровообращения и мочу мезобилиноген не попадает.
Основное
количество мезобилиногена из тонкой кишки поступает в толстую и здесь
восстанавливается до стеркобилиногена при участии анаэробной микрофлоры.
Образовавшийся стеркобилиноген в нижних отделах толстой кишки (в основном в
прямой кишке) окисляется до стерко-билина и выделяется с калом. Лишь небольшая
часть стеркобилиногена всасывается в систему нижней полой вены (попадает
сначала в геморроидальные вены) и в дальнейшем выводится с мочой.
Следовательно, в норме моча человека содержит следы стеркобилиногена (за сутки
его выделяется с мочой до 4 мг). К сожалению, до последнего времени в
клинической практике стеркобилиноген, содержащийся в нормальной моче,
продолжают называть уробилиногеном. На рис. схематично показаны пути
образования уробилиногеновых тел в организме
человека. В клинической практике укоренился термин «уробилиноген мочи». Под
этим термином следует понимать те производные билирубина
(билирубиноиды), которые обнаруживаются в моче. Положительная реакция на
уробилиноген может быть обусловлена повышенным содержанием того или иного
билирубиноида в моче и является, как правило, отражением патологии.
Определение
в клинике содержания билирубина
в крови (общего, непрямого и прямого), а также уробилиногена мочи имеет важное
значение при дифференциальной диагностике желтух различной этиологии (рис.
16.5). При гемолитической желтухе («надпеченочной») вследствие повышенного гемолиза
эритроцитов
и разрушения гемоглобина
происходит интенсивное образование непрямого билирубина
в ретикулоэндотелиальной системе (см. рис. 16.5, б). Печень оказывается
неспособной утилизировать такое большое количество непрямого билирубина,
что приводит к его накоплению в крови и тканях.
В печени при этом синтезируется повышенное количество прямого билирубина,
который с желчью попадает в кишечник. В тонкой кишке в повышенных количествах
образуется мезобилиноген и в последующем – стеркобилиноген. Всосавшаяся часть
мезобилиногена утилизируется печенью, а резорбирующийся в толстой кишке
стеркобилиноген выводится с мочой. Таким образом, для гемолитической желтухи в
типичных случаях характерны следующие клинико-лабораторные показатели:
повышение уровня общего и непрямого билирубина
в крови, в моче – отсутствие билирубина
(непрямой билирубин
не фильтруется почками) и положительная реакция на уробилиноген (за счет повышенного
попадания в кровь и мочу стеркобилиногена, а в тяжелых случаях – и за счет
мезобилиногена, не утилизирующегося печенью); лимонно-желтый оттенок кожных
покровов (сочетание желтухи и анемии); увеличение размеров селезенки; ярко
окрашенный кал.
Рис.
Патогенез билирубинемий при различны
хпатологических состояниях (схема). а - норма; б - гемолиз; в - застой в желчных капиллярах; г - поражение паренхиматозных клеток печени; 1 - кровеносный капилляр; 2 - клетки печени; 3 - желчный капилляр.
При
механической (обтурационной, или «подпеченочной») желтухе (см. рис. 16.5, в)
нарушен отток желчи (закупорка общего желчного протока камнем, рак
головки поджелудочной железы). Это приводит к деструктивным изменениям в печени
и попаданию элементов желчи (билирубин, холестерин,
желчные кислоты) в кровь. При полной обтурации общего желчного протока желчь не
попадает в кишечник, поэтому образования в кишечнике билирубиноидов не
происходит, кал обесцвечен и реакция на уробилиноген мочи отрицательная. Таким
образом, при механической желтухе в крови повышено количество общего билирубина
(за счет прямого), увеличено содержание холестерина
и желчных
кислот, а в моче – высокий уровень билирубина
(прямого). Клиническими особенностями обтурационной желтухи являются яркая
желтушная окраска кожи,
бесцветный кал, зуд кожи
(раздражение нервных окончаний желчными
кислотами, отлагающимися в коже). Следует заметить, что при длительно
сохраняющейся механической желтухе могут существенно нарушаться функции печени,
в том числе одна из главных – детоксикационная. В этом случае может произойти
частичный «отказ» печени от непрямого билирубина,
что может привести к его накоплению в крови. Иными словами, увеличение уровня
фракции непрямого билирубина
при механической желтухе является плохим прогностическим признаком.
При
паренхиматозной («печеночной») желтухе, возникающей чаще всего при ее вирусном
поражении, развиваются воспалительно-деструктивные процессы в печени, ведущие к
нарушению ее функций. На начальных этапах гепатита процесс захвата и
глюкуронирования непрямого билирубина
сохраняется, однако образующийся прямой билирубин
в условиях деструкции печеночной паренхимы
частично попадает в большой круг кровообращения, что ведет к желтухе. Экскреция
желчи также нарушена, билирубина
в кишечник попадает меньше, чем в норме. Меньше обычного образуется
мезобилиногена, и меньшее количество его всасывается в кишечнике. Однако даже
это небольшое количество поступающего в печень мезобилиногена не усваивается
ею. Мезобилиноген, «уклоняясь», попадает в кровь, а затем выделяется с мочой,
что предопределяет положительную реакцию на уробилиноген. Количество
образующегося стеркобилиногена также снижено, поэтому кал гипохоличный. Итак,
при паренхиматозной желтухе отмечается повышение в крови концентрации
общего билирубина,
преимущественно за счет прямого. В кале снижено содержание стеркобилиногена.
Реакция на уробилиноген мочи положительная за счет попадания в мочу
мезобилиногена. Следует отметить, что при прогрессирующем гепатите, когда
печень утрачивает свою детоксикационную функцию, в крови накапливается
значительное количество и непрямого билирубина.
Кроме того, при резко выраженном воспалении печени, ее «набухании», может
произойти сдавление желчных капилляров и протоков, возникнуть внутрипеченочный
холестаз, что придает паренхиматозной желтухе черты механической с соответствующей
клинико-лабораторной картиной (ахоличный кал, отсутствие реакции на
уробилиноген).
В
табл. приведены наиболее характерные сдвиги клинико-лабо-раторных показателей
при различных типах желтух.
Следует
иметь в виду, что в практике редко наблюдается желтуха какого-либо одного типа
в «чистом» виде. Чаще встречается сочетание того или иного типа. Так, при
выраженном гемолизе
неизбежно страдают различные органы, в том числе и печень, что может привнести
элементы паренхиматозной желтухи при гемолизе.
В свою очередь паренхиматозная желтуха, как правило, включает в себя элементы механической.
При механической желтухе, возникающей вследствие сдавливания большого сосочка
двенадцатиперстной кишки (фатерова соска) при раке
головки поджелудочной железы, неизбежен повышенный гемолиз
как следствие раковой интоксикации.
Желчь
– жидкий секрет желтовато-коричневого цвета, отделяется печеночными клетками.
В сутки у человека образуется 500–700 мл желчи (10 мл на 1 кг массы тела).
Желчеобразование происходит непрерывно, хотя интенсивность этого процесса на
протяжении суток резко колеблется. Вне пищеварения печеночная желчь переходит в
желчный пузырь, где происходит ее сгущение
в результате всасывания воды
и электролитов.
Относительная плотность печеночной желчи 1,01, а пузырной – 1,04. Концентрация
основных компонентов в пузырной желчи в 5–10 раз выше, чем в печеночной (табл.
16.3).
Предполагают,
что образование желчи начинается с активной секреции
гепатоцитами воды,
желчных
кислот и билирубина,
в результате которой в желчных канальцах появляется так называемая первичная
желчь. Последняя, проходя по желчным ходам, вступает в контакт с плазмой
крови, вследствие чего между желчью и плазмой
устанавливается равновесие электролитов,
т.е. в образовании желчи принимают участие в основном два механизма –
фильтрация и секреция.
В
печеночной желчи можно выделить две группы веществ.
Первая группа – это вещества,
которые присутствуют в желчи в количествах, мало отличающихся от их концентрации
в плазме
крови (например, ионы
Na+, К+, креатин и др.), что в какой-то мере служит
доказательством наличия фильтрационного механизма. Ко второй группе относятся
соединения, концентрация
которых в печеночной желчи во много раз превышает их содержание в плазме
крови (билирубин, желчные
кислоты и др.), что свидетельствует о наличии секреторного механизма. В
последнее время появляется все больше данных о преимущественной роли активной секреции
в механизме желчеобразования. Кроме того, в желчи обнаружен ряд ферментов,
из которых особо следует отметить щелочную фосфатазу
печеночного происхождения. При нарушении оттока желчи активность
данного фермента
в сыворотке
крови возрастает.
Основные
функции желчи. Эмульсификация. Соли желчных
кислот обладают способностью
значительно уменьшать поверхностное
натяжение. Благодаря этому они
осуществляют эмульгирование жиров в кишечнике, растворяют жирные кислоты и
нерастворимые в воде мыла. Нейтрализация кислоты. Желчь, рН которой немногим
более 7,0, нейтрализует кислый химус, поступающий из желудка, подготавливая его
для переваривания в кишечнике. Экскреция. Желчь – важный носитель экскрети-руемых желчных
кислот и холестерина. Кроме того, она удаляет из организма многие лекарственные вещества, токсины, желчные
пигменты и различные
неорганические вещества, такие, как медь, цинк и ртуть. Растворение холестерина. Как отмечалось, холестерин, подобно высшим
жирным кислотам, представляет
собой нерастворимое в воде соединение, которое сохраняется в желчи в
растворенном состоянии лишь благодаря присутствию в ней солей желчных
кислот и фосфатидилхолина. При недостатке желчных
кислот холестерин выпадает в осадок, при этом могут образовываться
камни. Обычно камни имеют окрашенное желчным
пигментом внутреннее ядро,
состоящее из белка. Чаще всего встречаются камни, у которых ядро
окружено чередующимися слоями холестерина и билирубината кальция. Такие камни содержат до 80% холестерина. Интенсивное образование камней отмечается при
застое желчи и наличии инфекции. При застое желчи встречаются камни, содержащие
90–95% холестерина, а при инфекции могут образовываться камни,
состоящие из билирубината кальция. Принято считать, что присутствие бактерий
сопровождается увеличением β-глюкуронидазной активности желчи, что приводит к расщеплению конъюгатов билирубина; освобождающийся билирубин служит субстратом для образования камней.
Продолжительность
жизни эритроцитов
составляет 120 дней, затем они разрушаются и освобождается гемоглобин.
Главными органами, в которых происходят разрушение эритроцитов
и распад гемоглобина,
являются печень, селезенка и костный мозг, хотя в принципе оба процесса могут
происходить и в клетках
других органов. Распад гемоглобина
в печени начинается с разрыва α-метиновой связи между I и II кольцами
порфиринового кольца. Этот процесс катализируется НАДФ-содержащей оксидазой и
приводит к образованию зеленого пигмента
вердоглобина (холеглобина):
В
приведенных структурных формулах здесь и далее в желчных
пигментах М – метильная СН3-группа, В – (—СН=СН2)
– винильная группа и П – (—СН2—СН2—СООН) – остаток
пропионовой кислоты.
Как видно из приведенных формул, в молекуле
вердоглобина еще сохраняются атом
железа
и белковый компонент. Имеются экспериментальные доказательства, что в этом
окислительном превращении гемоглобина
принимают участие витамин
С, ионы
Fe2+и другие кофакторы. Дальнейший распад вердоглобина, вероятнее
всего, происходит спонтанно с освобождением железа,
белка-глобина и образованием одного из желчных
пигментов – биливердина. Спонтанный распад сопровождается
перераспределением двойных
связей и атомов
водорода
в пиррольных кольцах и метиновых мостиках. Образовавшийся биливердин
ферментативным путем восстанавливается в печени в билирубин,
являющийся основным желчным
пигментом у человека и плотоядных животных:
Основное
место образования билирубина
– печень, селезенка и, по-видимому, эритроциты
(при распаде их иногда разрывается одна из метиновых связей в протопорфирине).
Образовавшийся во всех этих клетках
билирубин
поступает в печень, откуда вместе с желчью попадает в желчный пузырь. Билирубин,
образовавшийся в клетках
системы макрофагов, называется свободным, или непрямым, билирубином,
поскольку вследствие плохой растворимости
в воде
он легко адсорбируется на белках
плазмы
крови и для его определения в крови необходимо предварительное осаждение
белков
спиртом.
После этого билирубин
вступает во взаимодействие с диазореактивом Эрлиха.
В
крови взрослого здорового человека содержится относительно постоянное
количество общего билирубина
– от 4 до 26 мкмоль/л, в среднем 15 мкмоль/л. Около 75% этого количества
приходится на долю непрямого билирубина.
Повышение его концентрации
в крови до 35 мкмоль/л приводит к желтухе. Более высокий уровень билирубина
в крови вызывает явления тяжелого отравления. Непрямой билирубин,
поступая с током крови в печень, подвергается обезвреживанию путем связывания с
глюку-роновой кислотой. В этом процессе принимают участие особый фермент
УДФ-глюкуронилтрансфераза и УДФ-глюкуроновая кислота, являющаяся донором
глюкуроновой
кислоты. При этом к билирубину
присоединяются 2 остатка глюкуроновой
кислоты с образованием сравнительно индифферентного комплекса –
билирубин-диглюкуронида, хорошо растворимого в воде
и дающего прямую реакцию с диазореактивом. В желчи всегда присутствует прямой билирубин.
В крови количество прямого и непрямого билирубина,
а также соотношение между ними резко меняются при поражениях печени, селезенки,
костного мозга, болезнях крови и т.д., поэтому определение содержания обеих
форм билирубина
в крови имеет существенное значение при дифференциальной диагностике различных
форм желтухи. При желчнокаменной болезни в составе желчных камней наряду с
основным их компонентом – холестерином
всегда обнаруживается непрямой билирубин.
Вследствие плохой растворимости
в воде
он выпадает в осадок в желчном пузыре в виде билирубината кальция,
участвующего в формировании камней.
Дальнейшая
судьба желчных
пигментов, точнее билирубина,
связана с их превращениями в кишечнике под действием бактерий. Сначала
глюкуроновая кислота отщепляется от комплекса с билирубином
и освободившийся билирубин
подвергается восстановлению
в стеркобилиноген, который выводится из кишечника. В сутки человек выделяет
около 300 мг стеркобилиногена. Последний легко окисляется под действием света и
воздуха
в стеркобилин. Механизм бактериальных превращений билирубина
до стеркобилина до конца еще не расшифрован. Имеются данные, что промежуточными
продуктами восстановления
являются последовательно мезобилирубин и мезобилиноген (уробилиноген). После
всасывания небольшая часть мезобилиногена поступает через воротную вену в
печень, где подвергается разрушению с образованием моно- и дипиррольных
соединений. Кроме того, очень небольшая часть стеркобилиногена после всасывания
через систему геморроидальных вен попадает в большой круг кровообращения, минуя
печень, и в таком виде выводится с мочой. Однако называть его уробилиногеном не
совсем точно. Суточное содержание стеркобилиногена в моче составляет около 4
мг, и, пожалуй, именно стеркобилиноген является нормальной органической
составной частью мочи. Если с мочой выделяется повышенное содержание
уробилиногена (точнее, мезобилиногена), то это является свидетельством
недостаточности функции печени, например, при печеночной или гемолитической
желтухе, когда печень частично теряет способность извлекать этот пигмент
из крови воротной вены. Химически уробилиноген (мезобилиноген) неидентичен
стеркобилиногену (уробилиногену) мочи. Исчезновение стеркобилиногена
(уробилиногена) из мочи при наличии билирубина
и биливердина является свидетельством полного прекращения поступления желчи в
кишечник. Такое состояние часто наблюдается при закупорке протока желчного
пузыря (желчнокаменная болезнь) или общего желчного протока (желчнокаменная
болезнь, раковые поражения поджелудочной железы и др.).
Таким
образом, количественный и качественный
анализ желчных
пигментов в моче может представлять большой клинический интерес.
При поражении паренхимы печени (паренхиматозной,
или печеночной желтухе) у больных с гепатитами, циррозом, раком и другими
заболеваниями печени происходит повреждение ранее описанных четырех процессов,
протекающих в гепатоците.
http://www.youtube.com/watch?v=nXRWkorYFXc
Нарушение
захвата свободного билирубина печеночной клеткой (I) и связывания его с
глюкуроновой кислотой (II) ведет к увеличению в крови свободного (непрямого)
билирубина (Бн). Нарушение выделения билирубин-глюкуронида (прямого
билирубина) из печеночной клетки в желчные капилляры, обусловленное
воспалением, деструкцией, некрозами и снижением проницаемости мембран
гепатоцитов, приводит к регургитации желчи обратно в синусоиды и в общий кровоток
и, соответственно, к увеличению содержания в крови связанного (прямого)
билирубина (Бп).
Наконец,
нарушение функции гепатоцитов сопровождается также утратой способности
печеночной клетки захватывать и метаболизировать всосавшийся в кишечнике
уробилиноген (мезобилиноген), который в больших количествах попадает в
общий кровоток и выделяется с мочой в виде уробилина.
Таким
образом, при паренхиматозной желтухе в крови увеличено содержание
как свободного (непрямого), так и связанного (прямого) билирубина. Последний,
являясь хорошо растворимым в воде соединением, легко проходит почечный барьер и
появляется в моче, обуславливая ее темную окраску («цвет пива»). В моче также в
больших количествах присутствует уробилин (мезобилиноген). В кале содержание
стеркобилина может быть несколько уменьшено в связи с нарушением выделения
гепатоцитами желчи.
Механическая
(обтурационная) желтуха развивается при
обтурации внепеченочных желчевыводящих путей камнем или сдавлении общего
желчного протока опухолью (рак головки поджелудочной железы, метастазы рака в
лимфатические узлы ворот печени). В результате этого блокируется выделение
желчи в кишечник и, соответственно, не образуется уробилиноген (мезобилиноген и
стеркобилиноген). В связи с этим уробилин в моче и стеркобилин в кале полностью
отсутствуют (ахоличный кал). В крови значительно нарастает уровень связанного
(прямого) билирубина, поскольку его образование печеночной клеткой
длительное время не нарушено. Соответственно, в моче появляется большое
количество связанного билирубина и моча приобретает темный цвет («цвет пива»).
При
гемолитической желтухе происходит образование в РЭС большого количества свободного
(непрямого) билирубина, который полностью не успевает метаболизироваться в
печени, хотя функция гепатоцитов не нарушена и они работают с повышенной
нагрузкой. В результате в крови увеличивается содержание свободного
(непрямого) билирубина, который не проходит почечный барьер и не попадает в
мочу. Поскольку количество связанного (прямого) билирубина, выделяемого печенью
в кишечник (и, соответственно, стеркобилиногена) существенно увеличивается, в
моче значительно повышен уровень стеркобилиногена (уробилина),
попадающего в общий кровоток из геморроидальных вен.
Основные
лабораторные признаки желтух различного происхождения
Следует
иметь в виду, что иногда в клинической практике характер нарушений пигментного
обмена может существенно отличаться от приведенных выше лабораторных критериев
различных видов желтух. Так, при тяжелом поражении паренхимы печени, особенно в
сочетании с выраженным холестазом, выделение связанного (прямого) билирубина в
кишечник и, соответственно, образование метаболитов билирубина (мезобилиногена,
стеркобилиногена), резко уменьшено. Это приводит к значительному уменьшению
содержания стеркобилина в кале (ахоличный кал) и уробилина (мезобилиногена) в
моче. Наряду с резким возрастанием в крови уровня связанного (прямого)
билирубина, это может напоминать нарушения пигментного обмена, характерные для
механической желтухи.
Такая
ситуация нередко возникает при остром вирусном гепатите (болезни Боткина) и
других заболеваниях. Характерно, что по мере восстановления функции печени и
купирования холестаза в моче у таких больных появляется уробилин, а в кале —
стеркобилин. Соответственно выравниваются показатели связанного и несвязанного
билирубина в крови.
Нередко
при относительно длительном течении заболеваний, сопровождающихся закупоркой
желчного протока и развитием механической желтухи (желчнокаменная болезнь, рак
головки поджелудочной железы), в процесс вторично вовлекается также паренхима
печени, что приводит к частичному нарушению захватывания и связывания непрямого
билирубина и к повышению его содержания в сыворотке крови (наряду со связанным
билирубином, что характерно для механической желтухи). Одновременно изменяются
и другие лабораторные показатели, свойственные поражению паренхимы печени.
Обезвреживание
большинства ксенобиотиков происходит путём химической модификации и протекает в
2 фазы (рис. 12-1). В результате этой серии реакций ксенобиотики становятся
более гидрофильными и выделяются с мочой. Вещества, более гидрофобные или
обладающие большой молекулярной массой (>300 кД), чаще выводятся с жёлчью в
кишечник и затем удаляются с фекалиями.
Сисгема
обезвреживания включает множество разнообразных ферментов, под действием
которых практически любой ксенобиотик может быть модифицирован.
Микросомальные
ферменты катализируют реакции С-гидроксилирования, N-гидроксилирования, О-, N-,
S-дезалкилирования, окислительного дезаминирования, сульфоокисления и
эпоксидирования (табл. 12-1).
Таблица 12-1.
Возможные модификации ксенобиотиков в первой фазе обезвреживания
В мембранах
ЭР практически всех тканей локализована сисгема микросомального окисления (монооксигеназного
окисления). В эксперименте при выделении ЭР из клеток мембрана распадается на
части, каждая из которых образует замкнутый пузырёк - микросому, отсюда и
название - микросомальное окисление. Эта сисгема обеспечивает первую фазу
обезвреживания большинства гидрофобных веществ.
Рис. 12-1.
Метаболизм и выведение ксенобиотиков из организма. RH - ксенобиотик; К - группа, используемая при конъюгации (глутатион,
глюкуронил и др.); М - молекулярная масса. Из множества цитохром Р450 - зависимых реакций на рисунке
приведена только одна - схема гидроксилирования ксенобиотика. В ходе первой
фазы в структуру вещества RH вводится полярная группа ОН-. Далее происходит реакция конъюгации;
конъюгат в зависимости от растворимости и молекулярной массы удаляется либо
почками, либо с фекалиями.
В метаболизме
ксенобиотиков могут принимать участие ферменты почек, лёгких, кожи и ЖКТ, но
наиболее активны они в печени. К группе микросомальных ферментов относят
специфические оксидазы, различные гидролазы и ферменты конъюгации.
Вторая фаза -
реакции конъюгации, в результате которых чужеродное вещество, модифицированное
ферментными сисгемами ЭР, связывается с эндогенными субстратами - глюкуроновой
кислотой, серной кислотой, глицином, глутатионом. Образовавшийся конъюгат
удаляется из организма.
Микросомальные
оксидазы - ферменты, локализованные в мембранах гладкого ЭР, функционирующие в
комплексе с двумя внемитохондриальными ЦПЭ. Ферменты, катализирующие
восстановление одного атома молекулы О2 с образованием воды и
включение другого атома кислорода в окисляемое вещество, получили название
микросомальных оксидаз со смешанной функцией или микросомальных монооксигеназ.
Окисление с участием монооксигеназ обычно изучают, используя препараты
микросом.
1. Основные
ферменты микросомальных электронтранспортных цепей
Микросомальная
сисгема не содержит растворимых в цитозоле белковых компонентов, все ферменты -
мембранные белки, активные центры которых локализованы на цитоплазматической
поверхности ЭР. Сисгема включает несколько белков, составляющих
электронтранспортные цепи (ЦПЭ). В ЭР существуют две такие цепи, первая состоит
из двух ферментов - NADPH-P450 редуктазы и цитохрома Р450,
вторая включает фермент NADH-цитохром-b5 редуктазу, цитохром b5
и ещё один фермент - стеароил-КоА-десатуразу.
Электронтранспортная
цепь - NADPH-P450 редуктаза - цитохром Р450. В большинстве случаев донором электронов (e) для этой цепи служит NADPH,
окисляемый NАDРН-Р450 редуктазой. Фермент в качестве простетической группы содержит 2
кофермента - флавинаденинди-нуклеотид (FAD) и флавинмононуклеотид (FMN).
Протоны и электроны с NADPH переходят последовательно на коферменты NADPH-P450 редуктазы. Восстановленный FMN (FMNH2) окисляется цитохромом Р450 (см. схему).
Цитохром Р450 - гемопротеин, содержит простетическую группу гем и имеет участки
связывания для кислорода и субстрата (ксенобиотика). Название цитохром Р450 указывает на то, что максимум поглощения
комплекса цитохрома Р450 лежит в области 450 нм.
Окисляемый
субстрат (донор электронов) для NADH-цитохром b5 -редуктазы - NADH
(см. схему выше). Протоны и электроны с NADH переходят на кофермент редуктазы
FAD, следующим акцептором электронов служит Fe3+ цитохрома b5.
Цитохром b5 в некоторых случаях может быть донором электронов (e) для цитохрома Р450 или для
стеароил-КоА-десатуразы, которая катализирует образование двойных связей в
жирных кислотах, перенося электроны на кислород с образованием воды (рис.
12-2).
Рис. 12-2.
Электронтранспортные цепи ЭР. RH - субстрат цитохрома Р450; стрелками показаны реакции переноса
электронов. В одной системе NADPH окисляется NADPH цитохром Р450-редуктазой, которая затем передаёт
электроны на целое семейство цитохромов Р450. Вторая сисгема включает в себя окисление NADH цитохром b5-редуктазой, электроны переходят на
цитохром b5;
восстановленную форму цитохрома b5 окисляет стеароил-КоА-десатураза, которая переносит электроны на О2.
Один атом кислорода принимает 2 е и переходит в форму О2-. Донором электронов служит NADPH, который окисляется NADPH-цитохром Р450 редуктазой. О2- взаимодействует с протонами: О2- + 2Н+ → Н2О, и образуется вода. Второй атом
молекулы кислорода включается в субстрат RH, образуя гидроксильную группу
вещества R-OH.
NADH-цитохром
b5 редуктаза - двухдоменный
белок. Глобулярный цитозольный домен связывает простетическую группу -
кофермент FAD, а единственный гидрофобный "хвост" закрепляет белок в
мембране.
Цитохром b5- гемсодержащий белок, который имеет домен, локализованный на поверхности
мембраны ЭР, и короткий "заякоренный" в липидном бислое
спирализованный домен.
NADH-цитохром
b5 -редуктаза и цитохром b5, являясь
"заякоренными" белками, не фиксированы строго на определённых
участках мембраны ЭР и поэтому могут менять свою локализацию.
2.
Функционирование цитохрома Р450
Известно, что
молекулярный кислород в триплетном состоянии инертен и не способен
взаимодействовать с органическими соединениями. Чтобы сделать кислород
реакционно-способным, необходимо его превратить в синглетный, используя
ферментные системы его восстановления. К числу таковых принадлежит
моноксигеназная сисгема, содержащая цитохром Р450. Связывание в
активном центре цитохрома Р450 липофильного вещества RH и молекулы
кислорода повышает окислительную активность фермента.
Суммарное
уравнение реакции гидроксилирования вещества RH ферментами микросомального
окисления:
RH + О2
+ NADPH + Н+ → ROH + Н2О + NADP+ .
Субстратами Р450
могут быть многие гидрофобные вещества как экзогенного (лекарственные
препараты, ксенобиотики), так и эндогенного (стероиды, жирные кислоты и др.)
происхождения.
Таким
образом, в результате первой фазы обезвреживания с участием цитохрома Р450
происходит модификация веществ с образованием функциональных групп, повышающих
растворимость гидрофобного соединения. В результате модификации возможна потеря
молекулой её биологической активности или даже формирование более активного
соединения, чем вещество, из которого оно образовалось.
3. Свойства
системы микросомальногоокисления
Важнейшие
свойства ферментов микросомального окисления: широкая субстратная
специфичность, которая позволяет обезвреживать самые разнообразные по строению
вещества, и регуляция активности по механизму индукции.
Широкая
субстратная специфичность. Изоформы Р450
К настоящему
времени описано около 150 генов цитохрома Р450, кодирующих различные
изоформы фермента. Каждая из изоформ Р450 имеет много субстратов.
Этими субстратами могут быть как эндогенные липофильные вещества, модификация
которых входит в путь нормального метаболизма этих соединений, так и
гидрофобные ксенобиотики, в том числе лекарства. Определённые изоформы
цитохрома Р450 участвуют в метаболизме низкомолекулярных соединений,
таких как этанол и ацетон.
Рис. 12-3.
Транспорт электронов при монооксигеназном окислении с участием Р450. Связывание (1) в активном центре цитохрома Р450 вещества RH активирует
восстановление железа в теме - присоединяется первый электрон (2). Изменение
валентности железа увеличивает сродство комплекса P450-Fe2+·RH к молекуле кислорода (3).
Появление в центре связывания цитохрома Р450 молекулы О2
ускоряет присоединение второго электрона и образование комплекса P450-Fe2+O2--RH (4). На следующем этапе (5) Fe2+ окисляется, второй электрон
присоединяется к молекуле кислорода P450-Fe3+O22-. Восстановленный атом кислорода (О2-) связывает 2 протона, и образуется 1 молекула воды. Второй атом
кислорода идёт на построение ОН-группы (6). Модифицированное вещество R-OH
отделяется от фермента (7).
Регуляция
активности микросомальной системы окисления
Регуляция
активности микросомальной системы осуществляется на уровне транскрипции или
посттранскрипционных изменений. Индукция синтеза позволяет увеличить количество
ферментов в ответ на поступление или образование в организме веществ, выведение
которых невозможно без участия системы микросомального окисления.
В настоящее
время описано более 250 химических соединений, вызывающих индукцию
микросомальных ферментов. К числу этих индукторов относят барбитураты,
полициклические ароматические углеводороды, спирты, кетоны и некоторые
стероиды. Несмотря на разнообразие химического строения, все индукторы имеют
ряд общих признаков; их относят к числу липофильных соединений, и они служат
субстратами для цитохрома Р450.
Вторая фаза
обезвреживания веществ - реакции конъюгации, в ходе которых происходит
присоединение к функциональным группам, образующимся на первом этапе, других
молекул или групп эндогенного происхождения, увеличивающих гидрофильность и
умеНbшающих токсичность ксенобиотиков (табл. 12-2).
1. Участие
трансферам в реакциях конъюгации
Все ферменты,
функционирующие во второй фазе обезвреживания ксенобиотиков, относят к классу
трансфераз. Они характеризуются широкой субстратной специфичностью.
УДФ-глюкуронилтрансферазы
Локализированные
в основном в ЭР уридин-дифосфат (УДФ)-глюкуронилтрансферазы присоединяют
остаток глюкуроновой кислоты к молекуле вещества, образованного в ходе
мик-росомального окисления (рис.).
Рис.
Уридиндифосфоглюкуроновая кислота (УДФ-C6H9O6).
В общем виде
реакция с участием УДФ-глюкуронилтрансферазы записывается так:
ROH + УДФ-С6Н9О6
= RO-C6H9O6 + УДФ.
Сульфотрансферазы
Цитоплазматические
cульфотрансферазы катализируют реакцию конъюгации, в ходе которой остаток
серной кислоты (-SO3H) от 3'-фосфоаденозин-5'-фосфосульфата (ФАФС)
присоединяется к фенолам, спиртам или аминокислотам (рис. 12-5).
Рис.
3'-Фосфоаденозин-5'-фосфосульфат(ФАФ-SО3Н).
Таблица 12-2.
Основные ферменты и метаболиты, участвующие в конъюгации
Реакция с
участием сульфотрансферазы в общем виде записывается так:
ROH + ФАФ-SO3H = RO-SO3H + ФАФ.
Ферменты сульфотрансферазы
и УДФ-глюкуронилтрансферазы участвуют в обезвреживании ксенобиотиков,
инактивации лекарств и эндогенных биологически активных соединений.
Глутатионтрансферазы
Особое место
среди ферментов, участвующих в обезвреживании ксенобиотиков, инактивации
нормальных метаболитов, лекарств, занимают глутатионтрансферазы (ГТ).
Глутатионтрансферазы функционируют во всех тканях и играют важную роль в
инактивации собственных метаболитов: некоторых стероидных гормонов,
простагландинов, билирубина, жёлчных кислот, продуктов ПОЛ.
Известно
множество изоформ ГТ с различной субстратной специфичностью. В клетке ГТ в
основном локализованы в цитозоле, но имеются варианты ферментов в ядре и
митохондриях. Для работы ГТ требуется глутатион (GSH) (рис. 12-6).
Глутатион - трипептид Глу-Цис-Гли (остаток глутаминовой кислоты присоединён к
цис-теину карбоксильной группой радикала).
ГТ обладают
широкой специфичностью к субстратам, общее количество которых превышает 3000.
ГТ связывают очень многие гидрофобные вещества и инактивируют их, но химической
модификации с участием глугатиона подвергаются только те, которые имеют
полярную группу. То есть субстратами служат вещества, которые, с одной стороны,
имеют электрофильный центр (например, ОН-группу), а с другой стороны -
гидрофобные зоны. Обезвреживание, т.е. химическая модификация ксенобиотиков с
участием ГТ, может осуществляться тремя различными способами:
·
путём
конъюгации субстрата R с глутатионом (GSH):
R + GSH →
GSRH,
·
в результате
нуклеофильного замещения:
RX + GSH →
GSR + НХ,
·
восстановления
органических пероксидов до спиртов:
R-HC-O-OH + 2
GSH → R-HC-OH + GSSG + H2O
·
В реакции:
ООН - гидропероксидная группа, GSSG - окисленный глутатион.
Рис. 12-6.
Глутатион (GSH).
Сисгема обезвреживания
с участием ГТ и глутатиона играет уникальную роль в формировании резистентности
организма к самым различным воздействиям и является наиболее важным защитным
механизмом клетки. В ходе биотрансформации некоторых ксенобиотиков под
действием ГТ образуются тиоэфиры (конъюгаты RSG), которые затем превращаются в
меркаптаны, среди которых обнаружены токсические продукты. Но конъюгаты GSH с
большинством ксенобиотиков менее реакционно-способны и более гидрофильны, чем
исходные вещества, а поэтому менее токсичны и легче выводятся из организма
(рис.).
Рис. Oбезвреживание 1-хлор, 2,4-динитробензола с участием
глутатиона.
ГТ своими
гидрофобными центрами могут нековалентно связывать огромное количество
липофильных соединений (физическое обезвреживание), предотвращая их внедрение в
липидный слой мембран и нарушение функций клетки. Поэтому ГТ иногда называют
внутриклеточным альбумином.
ГТ могут
ковалентно связывать ксенобиотики, являющиеся сильными электролитами.
Присоединение таких веществ - "самоубийство" для ГТ, но
дополнительный защитный механизм для клетки.
Ацетилтрансферазы,
метилтрансферазы
Ацетилтрансферазы
катализируют реакции конъюгации - переноса ацетильного остатка от ацетил-КоА на
азот группы -SO2NH2, например в составе сульфаниламидов.
Мембранные и цитоплазматические метилтрансферазы с участием SAM метилируют группы
-Р=О, -NH2 и SH-группы ксенобиотиков.
2. Роль
эпоксидгидролаз в образовании диолов
Во второй
фазе обезвреживания (реакции конъюгации) принимают участие и некоторые другие
ферменты. Эпоксидгидролаза (эпоксидгидратаза) присоединяет воду к эпоксидам
бензола, бензпирена и другим полициклическим углеводородам, образованным в ходе
первой фазы обезвреживания, и превращает их в диолы (рис.).
Рис.
Обезвреживание бензантрацена. Е1 - фермент микросомальной системы; Е2 - эпоксидгидратаза.
Эпоксиды,
образовавшиеся при микросомальном окислении, являются канцерогенами. Они
обладают высокой химической активностью и могут участвовать в реакциях неферментативного
алкилирования ДНК, РНК, белков. Химические модификации этих молекул могут
привести к перерождению нормальной клетки в опухолевую.
Аминокислоты,
невсосавшиеся в клетки кишечника, используются микрофлорой толстой кишки в
качестве питательных веществ. Ферменты бактерий расщепляют аминокислоты и
превращают их в амины, фенолы, индол, скатол, сероводород и другие ядовитые для
организма соединения. Этот процесс иногда называют гниением белков в кишечнике.
В основе гниения лежат реакции декарбоксилирования и дезаминирования
аминокислот.
Образование и
обезвреживание n-крезола и фенола
Под действием
ферментов бактерий из аминокислоты тирозина могут образовываться фенол и крезол
путём разрушения боковых цепей аминокислот микробами (рис.).
Рис.
Катаболизм тирозина под действием бактерий. E - бактериальные
ферменты.
Всосавшиеся
продукты по воротной вене поступают в печеНb, где обезвреживание фенола и крезола
может происходить путём конъюгации с сернокислотным остатком (ФАФС) или с
глюкуроновой кислотой в составе УДФ-глюкуроната. Реакции конъюгации фенола и
крезола с ФАФС катализирует фермент сульфотрансфераза (рис.).
Рис.
Конъюгация фенола и крезола с ФАФС. E -
сульфотрансфераза.
Конъюгация
глюкуроновых кислот с фенолом и крезолом происходит при участии фермента
УДФ-глюкуронилтрансферазы (рис.).
Рис. Участие
УДФ-глюкуронилтрансферазы в обезвреживании крезола и фенола. E - УДФ-глюкуронилтрансфераза.
Продукты
конъюгации хорошо растворимы в воде и выводятся с мочой через почки. Повышение
количества конъюгатов глюкуроновой кислоты с фенолом и крезолом обнаруживают в
моче при увеличении продуктов гниения белков в кишечнике.
Образование и
обезвреживание индола и скатола
В кишечнике
из аминокислоты триптофана микроорганизмы образуют индол и скатол. Бактерии
разрушают боковую цепь триптофана, оставляя нетронутой кольцевую структуру.
Индол
образуется в результате отщепления бактериями боковой цепи, возможно, в виде
серина или аланина (рис.).
Рис.
Катаболизм триптофана под действием бактерий. E - бактериальные
ферменты.
Скатол и индол
обезвреживаются в печени в 2 этапа. Сначала в результате микросомального
окисления они приобретают гидроксильную группу. Так, индол переходит в
индоксил, а затем вступает в реакцию конъюгации с ФАФС, образуя индоксилсерную
кислоту, калиевая соль которой получила название животного индикана (рис.).
Рис. Участие
сульфотрансферазы в обезвреживании индола. E -
сульфотрансфераза.
Обезвреживание
бензойной кислоты
Синтез
гиппуровой кислоты из бензойной кислоты и глицина протекает у человека и большинства
животных преимущественно в печени (рис.).
Рис.
Образование гиппуровой кислоты из бензойной кислоты и глицина. E-глицинтрансфераза
Скорость этой реакции отражает функциональное
состояние печени.
В клинической
практике используют определение скорости образования и выведения гиппуровой
кислоты после введения в организм ксенобиотика бензойной кислоты
(бензойнокислого натрия) - проба Квика.
В плазме
крови множество как эндогенных, так и экзогенных липофильных веществ
транспортируются альбумином и другими белками.
Альбумин - основной белок плазмы крови, связывающий различные гидрофобные
вещества. Он может функционировать в качестве белка-переносчика билирубина,
ксенобиотиков, лекарственных веществ.
Помимо
альбуминов, ксенобиотики могут транспортироваться по крови в составе
липопротеинов, а также в комплексе с кислым α1-гликопротеином.
Особенность этого гликопротеина состоит в том, что он является индуцируемым
белком, участвующим в ответной реакции организма на изменения, происходящие в
состоянии стресса, например, при инфаркте миокарда, воспалительных процессах;
его количество в плазме увеличивается наряду с другими протеинами. Связывая
ксенобиотики, кислый α1-гликопротеин инактивирует их и переносит
в печень, где комплекс с белком распадается, и чужеродные вещества
обезвреживаются и выводятся из организма.
Участие
Р-гликопротеина в выведении ксенобиотиков
Очень важный
механизм выведения из клетки гидрофобных ксенобиотиков - функционирование
Р-гликопротеина (транспортная АТФ-аза). Р-гликопротеин - фосфогликопротеин с
молекулярной массой 170 кД, присутствующий в плазматической мембране клеток
многих тканей, в частности почек и кишечника. Полипептидная цепь этого белка
содержит 1280 аминокислотных остатков, образуя 12 трансмембранных доменов и два
АТФ-связывающих центра (рис.).
Рис. Строение
Р-гликопротеина. Р-гликопротеин - интегральный белок,
имеющий 12 трансмембранных доменов, пронизывающих бислой цитоплазматической
мембраны. N- и С-концы белка обращены в цитозоль. Участки Р-гликопротеина на
наружной поверхности мембраны гликозипированы. Область между шестым и седьмым
доменами имеет центры для присоединения АТФ и аутофосфорилирования.
В норме его
функция состоит в экскреции ионов хлора и гидрофобных токсичных соединений из
клеток.
Когда
гидрофобное вещество (например, противоопухолевое лекарство) проникает в
клетку, то оно удаляется из неё Р-гликопротеином с затратой энергии (рис.).
Рис.
Функционирование Р-гликопротеина. Заштрихованный
овал - противоопухолевое лекарство (гидрофобное вещество).
Уменьшение
количества лекарства в клетке снижает эффективность его применения при
химиотерапии онкологических заболеваний.
Индукция
защитных систем
Многие
ферменты, участвующие в первой и второй фазе обезвреживания, - индуцируемые
белки. Ещё в древности царь Митридат знал, что если сисгематически принимать
небольшие дозы яда, можно избежать острого отравления. "Эффект
Митридата" основан на индукции определённых защитных систем.
В мембранах
ЭР печени цитохрома Р450 содержится больше (20%), чем других
мембрано-связанных ферментов. Лекарственное вещество фенобарбитал активирует
синтез цитохрома Р450, УДФ-глюкуронилтрансферазы и эпоксид
гидролазы. Например, у животных, которым вводили индуктор фенобарбитал,
увеличивается площадь мембран ЭР, которая достигает 90% всех мембранных структур
клетки, и, как следствие, - увеличение количества ферментов, участвующих в
обезвреживании ксенобиотиков или токсических веществ эндогенного происхождения.
При химиотерапии
злокачественных процессов начальная эффективность лекарства часто постепенно
падает. Более того, развивается множественная лекарственная устойчивость, т.е.
устойчивость не только к этому лечебному препарату, но и целому ряду других
лекарств. Это происходит потому, что противоопухолевые лекарства индуцируют
синтез Р-гликопротеина, глутатионтрансферазы и глутатиона. Использование
веществ, ингибирующих или активирующих синтез Р-гликопротеина, а также ферменты
синтеза глутатиона, повышает эффективность химиотерапии.
Металлы
являются индукторами синтеза глутатиона и низкомолекулярного белка
металлотионеина, имеющих SH-группы, способные связывать их. В результате
возрастает устойчивость клеток организма к ядам и лекарствам.
Повышение
количества глутатионтрансфераз увеличивает способность организма
приспосабливаться к возрастающему загрязнению внешней среды. Индукцией фермента
объясняют отсутствие антиканцерогенного эффекта при применении ряда
лекарственных веществ. Кроме того, индукторы синтеза глутатионтрансферазы -
нормальные метаболиты - половые гормоны, йодтиронины и кортизол. Катехол-амины
через аденилатциклазную систему фосфорилируют глутатионтрансферазу и повышают
её активность.
Ряд веществ, в
том числе и лекарств (например, тяжёлые металлы, полифенолы, S-алкилы
глутатиона, некоторые гербициды), ингибируют глутатионтрансферазу.













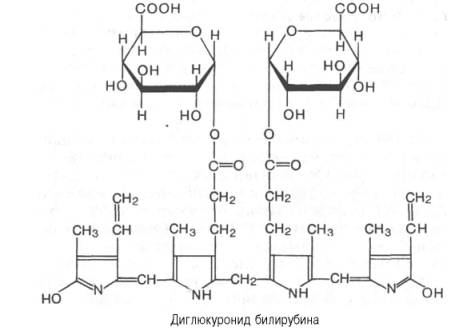{kind=link}
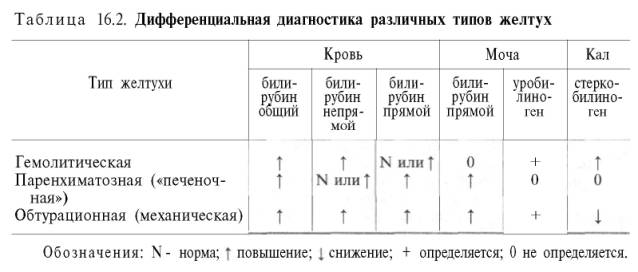{kind=link}
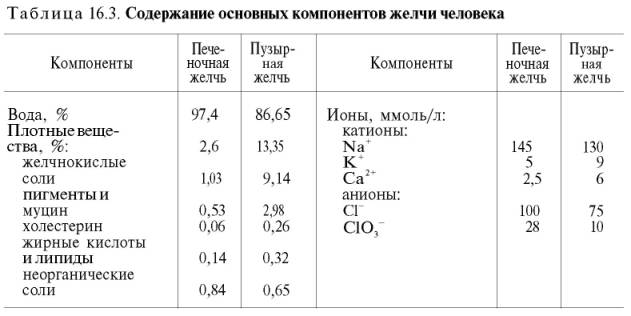{kind=link}
{kind=link}
{kind=link}
{kind=link}









{kind=link}