

Investigation of physico-chemical properties and
chemical composition of
Investigation of pathological components of urine.

Kidney – the couple organ, which is responsible for excriting of final products of metabolism and for homeostasis. They regulate water and mineral metabolism, acid-base balance, excriting of nitrogenous slags, osmotic pressure. Also they regulate arterial pressure and erhythropoesis.
Understanding how the urinary system helps maintain homeostasis by removing harmful substances from the blood and regulating water balance in the body is an important part of physiology. Your kidneys, which are the main part of the urinary system, are made up of millions of nephrons that act as individual filtering units and are complex structures themselves. The ureters, urethra, and urinary bladder complete this intricate system.
Kidney functions in organism:
a) excretion of final metabolic products;
b) maintaining of acid-base balance;
c) water-salts balance regulation;
d) endocrine function.(The kidneys secrete a variety of hormones, including erythropoietin, and the enzyme renin. Erythropoietin is released in response to hypoxia (low levels of oxygen at tissue level) in the renal circulation. It stimulates erythropoiesis (production of red blood cells) in the bone marrow. Calcitriol, the activated form of vitamin D, promotes intestinal absorption of calcium and the renalreabsorption of phosphate. Part of the renin-angiotensin-aldosterone system, renin is an enzyme involved in the regulation of aldosterone levels.)
The kidney participates in whole-body homeostasis, regulating acid-base balance, electrolyte concentrations, extracellular fluid volume, and regulation of blood pressure. The kidney accomplishes these homeostatic functions both independently and in concert with other organs, particularly those of the endocrine system. Various endocrine hormones coordinate these endocrine functions; these include renin, angiotensin II, aldosterone, antidiuretic hormone, and atrial natriuretic peptide, among others.
Many of the kidney's functions are
accomplished by relatively simple mechanisms of filtration, reabsorption, and
secretion, which take place in the nephron.
Filtration, which takes place at therenal
corpuscle, is the process by which cells and large proteins are
filtered from the blood to make an ultrafiltrate that eventually becomes urine.
The kidney generates
http://www.youtube.com/watch?v=6x5pVoMb_vI&feature=related
Physical and chemical characteristics and components of urine:
a) volume, physical and chemical properties of urine;
b) inorganic components of urine;
c) organic components of urine.
Key words and phrases:
http://www.youtube.com/watch?v=glu0dzK4dbU
Nephron – is the structural and functional unit of kidney.
Urine – fluid with different organic and inorganic compounds, which must be excreted (excess of water, final products of nitrogen metabolism, xenobiotics, products of protein’s decay, hormones, vitamins and their derivates). Most of them present in urine in a bigger amount than in blood plasma. So, urine formation – is not passive process (filtration and diffusion only).
In basis of urine formation lay 3 processes: filtration, reabsorbtion and secretion.
Glomerulal filtration. Water and low weight molecules go to the urine with help of following powers: blood hydrostatic pressure in glomerulas (near 70 mm Hg), oncotic press ure of blood plasma proteins (near 30 mm Hg) and hydrostatic pressure of plasma ultrafiltrate in glomerulal capsule (near 20 mm Hg). In normal conditions, as You see, effective filtration pressure is about 20 mm Hg.
Hydrostatic pressure depends from correlation between opening of a. afference and a. efference.
Primary urine formed in result of filtration (about 200 L per day). Between all blood plasma substances only proteins don’t present in primary urine. Most of these substances are undergone to the following reabsorbtion. Only urea, uric acid, creatinin, and other final products of different metabolic pathways aren’t undergone to the reabsorbtion.
For evaluate of filtration used clearance (clearance for some substance – it is an amount of blood plasma in ml, which is cleaned from this substance after 1 minute passing through kidney).
Drugs, which stimulate blood circulation in kidney (theophyllin), also stimulate filtration. Inflammatory processes of renal tissue (nephritis) reduce filtration, and azotaemia occurred (accumulation of urea, uric acid, creatinin, and other metabolic final products).
Glomerulal filtration.
Water and low weight molecules go to the urine with help of following powers:
blood hydrostatic pressure in glomerulas (near
Hydrostatic pressure depends from correlation between opening of a. afference and a. efference.
Primary urine formed in result of filtration (about
For evaluate of filtration used clearance (clearance for some substance – it is a amount of blood plasma in ml, which is cleaned from this substance after 1 minute passing through kidney).
Drugs which stimulate blood circulation in kidney (theophyllin), also stimulate filtration. Inflammatory processes of renal tissue (nephritis) reduce filtration, and azotaemia occurred (accumulation of urea, uric acid, creatinin, and other metabolic final products).
The development of the clearance concept
ABSTRACT: The clearance concept is central to three critical areas of nephrology; it was a key feature to early conceptual analysis of the nature of urine formation, it was utilized as a measure of kidney function in advancing renal diseases, and it was a pivotal concept to elucidate the physiology of the kidneys. This paper describes the clearance concept as currently understood and then it examines how clearance was utilized to understand these various aspects of kidney function.
Introduction
The concept of clearance is central to three major areas of nephrology. First, the nature of urine for mation was explored to a great extent using clearance techniques. Second, the early search for measures of kidney function with advancing disease resorted to clearance procedures, particularly involving urea and creatinine. Third, the physiology of the kidney was examined and developed with great power and sophistication by the deepening theoretical understanding of the concept of clearance accompanied by ingenious analytical techniques and procedures. There is an additional domain which hovered over the studies of urine formation. This had to do with the per vasive resort to vitalism as an explanation of physiologic regulation. My task was to examine the birth and evolution of the clearance concept. For purposes of exposition, it may be helpful to describe first the clearance concept as it is currently understood and then recount how the concept emerged and developed during the 19th and early 20th centuries, as it was repeatedly invoked to analyze the process of urine formation, the failure of renal function, and the nature of physiologic regulation.
THE CONCEPT OF CLEARANCE
Figure 1 gives the definition of clearance in currently conventional units of time and concentration. It is evident from the formula that the numerator is a rate of excretion (mg/mm); the denominator is a plasma concentration (mg/mL). Therefore, the clearance of any substance is expressed as mL/min (mg/min x mL/mg=mL/min). Clearance, therefore, has the dimensions of a volume per unit time. This simultaneous measurement of the excretion rate of a solute and a flow rate of fluid from which the solute is derived has resulted in some confusion. Fig 2, modified from Cassin and Vogh, emphasizes that the kidney removes (clears) a small fraction of a substance from each mL of total flow. The clearance, therefore, of any substance is the virtual volume of plasma flow required to supply the amount of the substance excreted in any one minute.
NATURE OF URINE FORMATION
The
historical evolution of the clearance concept is intimately connected with
studies examining the nature of urine formation. Particularly noteworthy
reviews have been published by Smith (2), Bradley (3), Thurau, Davis and
Haberle (4), Gottschalk (5), Schuster and Seldin (14). In the early 19th centur
y, Johannes Muller (18011858) advanced a theor y of urine formation that rested
on two prevalent concepts current at the time: 1) fluid movement was a
secretory process mediated by glands; 2) the activity of the secretor y system
required vitalistic forces that could not be reduced to physical processes.
Despite enormous contributions to microscopic anatomy, he denied that the
glomerulus was directly connected with the renal tubules, and ascribed urine
formation to the secretory activity of the tubules, regarding the kidney as a
gland. Notwithstanding the powerful currents of vitalism at the time, Carl
Ludwig (1816-1895) came to the study of kidney function with an uncompromising
physicochemical orientation. He appreciated the role of the afferent and
efferent arteriole in elevating the hydrostatic pressure in the intervening
glomerulus, thereby facilitating the movement of a protein-free ultrafiltrate,
containing all the elements to be found in the urine, and restraining the
passage of protein and formed elements. To account for the different
composition of blood and urine, Ludwig proposed that some unspecified chemical
force promoted active sodium chloride reabsorption while some property of the
tubular wall restrained urea back-diffusion. No vital force was postulated,
although the nature of the “chemical force” promoting reabsorption was
unspecified. Simultaneously and independently, William Bowman (1816-1892) also
postulated that water was separated from blood at the glomerulus, but he
assumed that solutes remained in the blood and were subsequently secreted into
the urine by the tubules. This was an expression of the prevalent view of
glandular secretion mediating solute movement. The central feature of Ludwig’s
theory that urine formation was critically linked with glomerular pressure, was
challenged by Rudolph Heidenhain (1834-1897). On the basis of calculations of a
clearance type, he concluded that to attribute urea excretion to filtration
alone would require
His conception of a trifold process of ultrafiltration at the glomerulus by physical forces, reabsorption of most of the filtrate in part by active tubular processes, and active secretion of certain solutes by the tubular epithelia is remarkably close to modern views. His summary is worth quoting . It should be emphasized that the use of clearance calculations by Heidenhain and their reinterpretation by Cushny served to make creditable the conceptual model of the comparatively modest magnitude of urine flow in a setting of huge volumes of glomerular filtration.
DIGRESSION ON VITALISM
Although Cushny may have over-emphasized the commitment of Heidenhain and others to vitalism, there is no question that the concept of vital activity, not reducible to physical forces, was a powerful conceptual factor that infected the theories of renal function. For most of the 19th centur y, a basic problem in biology was conceived to be the distinction between living and non-living matter. A mechanistic explanation assumed that organic and non-organic matter were not irreducibly different. A vitalistic explanation assumed that a reduction of living to non-living phenomena is in principle impossible. Embryology was a dominating biologic discipline. To provide a flavor of the intellectual climate surrounding the study of renal function, it may be helpful to review briefly the prevailing embryologic studies. Landmark studies exemplified by the work of Hans Driesch are summarized in a comprehensive publication in 1914 (7). In a series of studies on embr yonic sea-urchins, he demonstrated that rearrangement of cells at the blastomere stage had no effect on normal development. Moreover, a single blastomere, isolated from the rest at the two - or fourcell stage, can develop into a normal sea-urchin embr yo.
The conclusion was drawn that spatio temporal location is irrelevant to development, and that non-physical forces “entelechies” are “wholemaking” factors which have no quantitative characteristics. It was only the gradual advancement of physical and biologic science that could meet the vitalistic arguments. Organic chemistr y was shown to be a misnomer. The synthesis of urea, heretofore found only in living organisms, from CO2 and NHby3 Wöhler in 1828 (7) led to the view that organic chemistr y was simply the chemistr y of carbon compounds. Purpose and purposiveness were explained by reference to integrated and adjustible feed-back systems. The ability of blastomere cells to develop differently in different transplant locations in ontogenesis, unlike a machine where each part fulfills a designed function, is explainable in principle by genetic theory. And finally “energy” input required to impart selectivity is not confined to hydrostatic or oncotic forces. On a conceptual level, it was pointed out by the logical positivist philosopher C.I. Hengel that vitalism has no predictive power, offering neither verifiable predictions nor providing models of coherent mechanisms. It was the increasing power of the physical sciences that gradually undermined the recourse to postulated entities which could not be identified, characterized, or worst of all, refuted. It was these reasons which led Ludwig and Cushny to vigorously reject vitalistic explanations.
UREA AND UREA CLEARANCE
Bright in 1836 recognized that the concentration of blood urea rose in patients with chronic renal disease (3). Ambard (8) showed that the blood level of urea was related to urea excretion and formulated an equation which was designed to register impairment of renal function.
However, the equation involved a square root function which obscured the physiologic significance of the relationship between urinar y excretion and blood urea concentration. Addis (9), in 1917, showed that at maximal urine flows the ratio of the excretion of urea per hour and the blood urea concentration was constant in any one individual. This expression represented the urea clearance per hour, an approximation of glomerular filtration rate. Austin, Stillman and Van Slyke (10) showed that the rate of urine flow influenced urea excretion independently of the blood level and renal excretory capacity. In a later study (11), it was demonstrated that above urine flows of 2ml/min (augmentation limit), the relationship between urea excretion and plasma concentration in any one individual was constant, and expressed by a simple formu1a:
Curea = Urea / Purea
The term, clearance, was introduced with this analysis. Since blood urea concentration is frequently used as an index of filtration rate, it is worthwhile examining the factors which influence it independently of intrinsic renal function. It has alredy been pointed out that urine flow influences urea clearance. Urea undergoes a complex intrarenal recycling process, the fractional reabsorption increasing from 35% of the filtered load in hydrated states to 60% in dehydration. The blood urea concentration is influenced by a variety of factors independent of renal function. Changes in urine flow affect blood urea in a manner which depends on the nephron segment where fluid is being reabsorbed.
Fig. 4
Fig. 5
The proximal tubule is highly permeable to urea, and is the principal segment of passive reabsorption. The distal nephron is less permeable to urea, even in the presence of antidiuretic hormone. If ever ything else is left constant, salt depletion will produce more azotemia at the same low rate of urine flow than will water restriction, because salt depletion accelerates proximal reabsorption while water restriction accelerates principally distal reabsorption (12). Protein loads also influence blood urea concentration independent of renal function. Figure 4 lists the sources of protein loads. Factors 1-5 serve to increase protein loads while factor 6 reduces it. Figure 5 (13) illustrates the effect of protein intake at various levels of renal function.
Fig. 6
It should be emphasized that at low filtration rates, the BUN is very sensitive to protein loads, as is illustrated by Figure 6 (13). Figure 7 summarizes the various factors which influence both the BUN and urea clearance. It is evident that the interpretation of the BUN as a rough index of GFR requires correction for the numerous factors which influence its concentration independent of renal function. Urea clearance circumvents some of the distorting effects of protein loads. Nevertheless, it is always less than inulin clearance, but tends to rise toward inulin clearance with advanced renal failure. These various factors are discussed in detail in ref. 14.
PHYSIOLOGY OF THE KIDNEY
Although Addis and Van Slyke had published landmark studies on urea clearance as a measure of renal function, the precise relation between urea clearance and glomerular filtration rate was not appreciated. Rehberg (15, 16) introduced creatinine as a marker of glomerular filtration rate, but was unaware that it was secreted by the tubules, and therefore would give falsely high values, especially if its plasma concentration was raised by infusions. Smith was skeptical that creatinine would be an ideal marker for glomerular filtration, since it underwent secretion in aglomerular fish, and might do the same in mammals, a supposition that proved correct. He then went on to elaborate the criteria for an ideal marker of GFR (Fig. 8).
Fig. 7
Fig. 8
The failure of sugars to be secreted in aglomerular fish led Smith ultimately to identify inulin as an ideal marker (17, 18). Simultaneously and independently, Richards and his associates also demonstrated in micropuncture studies that inulin fulfilled the requirements for an ideal marker of GFR (19). In Figure 9, inulin excretion is shown to increase in direct proportion to its plasma concentration when GFR is constant (a); inulin clearance is constant over a wide range of plasma inulin concentrations (b); inulin clearance is constant over a wide range of urine flows (c). Findings such as these established inulin as a kind of gold standard for GFR (14). Smith went on to develop methods for measuring renal blood flow, utilizing diodrast as a marker and the Fick principle to calculate total renal blood flow. Since the Fick principle required renal vein catheterization, it was unsuitable for routine use. Para-amino hippurate (PAH) was identified as a substance which, at low plasma concentrations, was almost completely secreted into the tubular urine. This eliminated the need for renal catherization, since renal venous PAH could be assumed to be close to zero. Subsequent studies of tubular maximum transport capacity using glucose and many other substances provided a measure of functioning tubular mass.
Fig. 9
Fig. 10
These various measures allowed Smith to portray the various functional aspects of normal and diseased kidneys in remarkable detail, as summarized in his Porter lectures (20). From a clinical and physiologic point of view, the various measures of renal function that Smith and others explored over the years are summarized in Figure 10. Smith has remarked how fruitful the clearance concept has proved to be. From rather tentative beginnings it has stimulated the search for novel analytic procedures, allowed for the assessment of renal function, and most of all provided a conceptual rallying point for insight and understanding of renal physiology.
Reabsorption. Lenght of renal tubules is
about
All substances can be divided into 3 group:
1. Actively reabsorbed substances.
2. Substances, which are reabsorbed in a little amount.
3. Non-reabsorbed substances.
To the first group belong Na+, Cl-, Mg2+, Ca2+, H2O, glucose and other monosaccharides, amino acids, inorganic phosphates, hydrocarbonates, low-weight proteins, etc.
Na+ reabsorbed by active transport to the epitelium cell, then – into the extracellular matrix. Cl- and HCO3- following Na+ according to the electroneutrality principle, water – according to the osmotic gradient. From extracellular matrix substances go to the blood vessels. Mg2+ and Ca2+ are reabsorbed with help of special transport ATPases. Glucose and amino acids use the energy of Na+ gradient and special carriers. Proteins are reabsorbed by endocytosis.
Urea and uric acid are little reabsorbable substances.
Creatinin, mannitol, inulin and some other substances are non-reabsorbable.
Henle’s loop play important role in the reabsobtion process. Its descendent and ascendent parts create anti-stream system, which has big capacity for urine concentration and dilution. Fluid which passes from proximal part of renal tubule to the descendent part of Henle’s loop, where concentration of osmotic active substances higher than in kidney cortex. This concentration is due to activity of thick ascendent part of Henle’s loop, which is non-penetrated for water and which cells transport Na+ and Cl- into the interstitium. Wall of descendent part is penetrated for water and here water pass into the interstitium by osmotic gradient but osmotic active substances stay in the tubule. Ascendent part continue to reabsorb salt hypertonically, even in the absence of aldosteron, so that fluid entering the distal tubule still has a much lower osmolality than does interstitial fluid.
Extracellular homeostasis
The kidney is responsible for maintaining a balance of the following substances:
|
Substance |
Description |
||||
|
If glucose is not reabsorbed by the kidney, it appears in the urine, in a condition known as glycosuria. This is associated with diabetes mellitus. |
reabsorption (almost 100%) viasodium-glucose transport proteins(apical) and GLUT (basolateral). |
– |
– |
– |
|
|
All are reabsorbed nearly completely. |
reabsorption |
– |
– |
– |
|
|
Regulation of osmolality. Varies with ADH |
reabsorption (50%) via passive transport |
secretion |
– |
reabsorption in medullary collecting ducts |
|
|
Uses Na-H antiport, Na-glucose symport, sodium ion channels (minor) |
reabsorption (65%, isosmotic) |
reabsorption (25%, thick ascending, Na-K-2Cl symporter) |
reabsorption (5%, sodium-chloride symporter |
reabsorption (5%, principal cells), stimulated by aldosterone via ENaC |
|
|
Usually follows sodium. Active (transcellular) and passive (paracellular) |
reabsorption |
reabsorption (thin ascending, thick ascending, Na-K-2Cl symporter) |
reabsorption (sodium-chloride symporter) |
– |
|
|
absorbed osmotically along with solutes |
reabsorption (descending) |
– |
reabsorption (regulated by ADH, via arginine vasopressin receptor 2) |
||
|
Helps maintain acid-base balance. |
reabsorption (80–90%) |
reabsorption (thick ascending) |
– |
||
|
Uses vacuolar H+ATPase |
– |
– |
– |
secretion (intercalated cells) |
|
|
Varies upon dietary needs. |
reabsorption (65%) |
reabsorption (20%, thick ascending, Na-K-2Cl symporter) |
– |
secretion (common, via Na+/K+-ATPase, increased by aldosterone), or reabsorption (rare, hydrogen potassium ATPase) |
|
|
reabsorption |
reabsorption (thick ascending) via passive transport |
– |
– |
||
|
Calcium and magnesium compete, and an excess of one can lead to excretion of the other. |
reabsorption |
reabsorption (thick ascending) |
reabsorption |
– |
|
|
Excreted as titratable acid. |
reabsorption (85%) viasodium/phosphate cotransporter. inhibited by parathyroid hormone. |
– |
– |
– |
|
|
|
reabsorption (100%) viacarboxylate transporters. |
– |
– |
– |
The body is very sensitive to its pH. Outside the range of pH that is compatible with life, proteins are denatured and digested, enzymes lose their ability to function, and the body is unable to sustain itself. The kidneys maintain acid-base homeostasis by regulating the pH of the blood plasma. Gains and losses of acid and base must be balanced. Acids are divided into "volatile acids" and "nonvolatile acids".[14] See also titratable acid.
The major homeostatic control point for maintaining this stable balance is renal excretion. The kidney is directed to excrete or retain sodium via the action of aldosterone, antidiuretic hormone(ADH, or vasopressin), atrial natriuretic peptide (ANP), and other hormones. Abnormal ranges of the fractional excretion of sodium can imply acute tubular necrosis or glomerular dysfunction.
Some substances (K+, ammonia and other) are secreted into urine in the distal part of tubules. K+ is changed to Na+ by the activity of Na+-K+ATPase.
WATER REGULATION BY THE KIDNEYS
The water content of the body can vary depending on
various factors. Hot weather and physical activity such as exercise make us
sweat and so lose body fluids. Drinking tends to be at irregular intervals when
socially convenient. This means that sometimes the body has too little water
and needs to conserve it and sometimes too much water and needs to get rid of
it. Most of the control of water conservation takes place in the distal and
collecting tubules of the nephrons under control of anti-diuretic hormone,
(ADH), sometimes called vasopressin. This hormone is released by the posterior
pituitary under control of the hypothalamus in the mid-brain area. The
hypothalamus monitors the water content of the blood. If the blood contains too
little water (indicating dehydration) then more ADH is released. If the blood stream contains too much water (indicating over-hydration)
then less ADH is released into the blood
Release of ADH from the posterior pituitary into the blood stream
ADH released from the pituitary travels in the blood stream to the peritubular capillaries of the nephron. ADH binds to receptors on the distal and collecting tubules of the nephrons which causes water channels to open in the tubule walls. This allows water to diffuse through the tubule walls into the interstitial fluid where it is collected by the peritubular capillaries. The more ADH present, the more water channels are open and the more water is reabsorbed,
Reabsorption of water from the filtrate under the influence of ADH
Over 99% of the filtrate produced each day can be reabsorbed. The amount of water reabsorbed from the filtrate back into the blood depends on the water situation in the body. When the body is dehydrated, most of the filtrate is reabsorbed but note that even in cases of extreme of water shortage, the kidneys will continue to produce around 500 ml of urine each day in order to perform their excretory function.
Peculiarities of biochemical processes in kidney.
Kidney have a very high level of metabolic processes. They use about 10 % of all O2, which used in organism. During 24 hours through kidney pass 700-900 L of blood. Carbohydrates are the main fuel for kidney. Glycolysis, ketolysis, aerobic oxidation and phosphorylation are very intensive in kidney. A lot of ATP formed in result.
Metabolism of proteins also presents in kidney in high level. Especially, glutamine deaminase is very active and a lot of free ammonia formed. In kidney take place the first reaction of creatin synthesis.
Kidney have plenty of different enzymes: LDG (1, 2, 3, 5), AsAT, AlAT. Specific for kidney is alanine amino peptidase, 3rd isoform.
Utilization of glucose in cortex and medulla is differs one from another. Dominative type of glycolysis in cortex is aerobic way and CO2 formed in result. In medulla dominative type is anaerobic and glucose converted to lactate.
Two sources contribute to the renal ammonia: blood ammonia (is about one-third of excreted ammonia), and ammonia formed in the kidney. The predominant source for ammonia production within the kidney is glutamine, the most abundant amino acid in plasma, but a small amount may originate from the metabolism of other amino acids such as asparagine, alanine, and histidine. Ammonia is secreted into the tubular lumen throughout the entire length of the nephron. Secretion occurs both during normal acid-base balance and in chronic acidosis. Metabolic acidosis is accompanied by an adaptive increase in renal ammonia production with a corresponding increase in urinary ammonium excretion.
Kidney cortex like liver appear to be unique in that it possess the enzymatic potential for both glucose synthesis from noncarbohydrate precursors (gluconeogenesis) and glucose degradation via the glycolytic pathway. Gluconeogenesis is important when the dietary supply of glucose does not satisfy the metabolic demands. Under these conditions, glucose is required by the central nervous system, the red blood cells, and, possibly, other tissues which cannot obtain all their energy requirements from fatty acids or ketone body oxidation. Also, gluconeogenesis may be important in the removal of excessive quantities of glucose precursors from the blood (lactate acid after severe exercise for example). Although the biosynthetic pathways are similar, there are several important differences in the factors, which regulate gluconeogenesis in the two organs. 1) The liver utilizes predominately pyruvate, lactate and alanine. The kidney cortex utilizes pyruvate, lactate, citrate, α-ketoglutarate, glycine and glutamine. 2) Hydrogen ion activity has little effect upon hepatic gluconeogenesis, but it has marked effects upon renal gluconeogenesis. Thus, when intracellular fluid pH is reduced (metabolic acidosis, respiratory acidosis or potassium depletion), the rates of gluconeogenesis in slices of renal cortex are markedly increased. The ability of the kidney to convert certain organic acids to glucose, a neutral substance, is an example of a nonexcretory mechanism in the kidney for pH regulation.
Renin-Angiotensin mechanism
The renin–angiotensin system (RAS) or the renin–angiotensin–aldosterone system (RAAS) is a hormone system that regulates blood pressure and water (fluid) balance.
When blood volume is low, juxtaglomerular cells in the kidneys secrete renin directly into circulation. Plasma renin then carries out the conversion of angiotensinogen released by the liver to angiotensin I. Angiotensin I is subsequently converted to angiotensin II by the enzyme angiotensin converting enzyme found in the lungs. Angiotensin II is a potent vaso-active peptide that causes blood vessels to constrict, resulting in increased blood pressure. Angiotensin II also stimulates the secretion of the hormone aldosterone from the adrenal cortex. Aldosterone causes the tubules of the kidneys to increase the reabsorption of sodium and water into the blood. This increases the volume of fluid in the body, which also increases blood pressure.
If the renin–angiotensin–aldosterone system is abnormally active, blood pressure will be too high. There are many drugs that interrupt different steps in this system to lower blood pressure. These drugs are one of the main ways to control high blood pressure (hypertension), heart failure,kidney failure, and harmful effects of diabetes.
The system can be activated when there is a loss of blood volume or a drop in blood pressure (such as in hemorrhage). This loss of pressure is interpreted by baroreceptors in the carotid sinus. In alternative fashion, a decrease in the filtrate NaCl concentration and/or decreased filtrate flow rate will stimulate the macula densa to signal the juxtaglomerular cells to release renin.
· If the perfusion of the juxtaglomerular apparatus in the kidney's macula densa decreases, then the juxtaglomerular cells (granular cells, modified pericytes in the glomerular capillary) release the enzyme renin.
· Renin cleaves a zymogen, an inactive peptide, called angiotensinogen, converting it into angiotensin I.
· Angiotensin I is then converted to angiotensin II by angiotensin-converting enzyme (ACE),[5] which is thought to be found mainly in lung capillaries. One study in 1992 found ACE in all blood vessel endothelial cells.[6]
· Angiotensin II is the major bioactive product of the renin-angiotensin system, binding to receptors on intraglomerular mesangial cells, causing these cells to contract along with the blood vessels surrounding them and causing the release of aldosterone from the zona glomerulosa in the adrenal cortex. Angiotensin II acts as an endocrine, autocrine/paracrine,and intracrinehormo.
Cardiovascular effects

It is believed that angiotensin I may have some minor activity, but angiotensin II is the major bio-active product. Angiotensin II has a variety of effects on the body:
· Throughout the body, it is a potent vasoconstrictor of arterioles.
· In the kidneys, it constricts glomerular arterioles, having a greater effect on efferent arterioles than afferent. As with most other capillary beds in the body, the constriction of afferent arterioles increases the arteriolar resistance, raising systemic arterial blood pressure and decreasing the blood flow. However, the kidneys must continue to filter enough blood despite this drop in blood flow, necessitating mechanisms to keep glomerular blood pressure up. To do this, angiotensin II constricts efferent arterioles, which forces blood to build up in the glomerulus, increasing glomerular pressure. The glomerular filtration rate(GFR) is thus maintained, and blood filtration can continue despite lowered overall kidney blood flow. Because the filtration fraction has increased, there is less plasma fluid in the downstream peritubular capillaries. This in turn leads to a decreased hydrostatic pressure and increased oncotic pressure (due to unfiltered plasma proteins) in the peritubular capillaries. The effect of decreased hydrostatic pressure and increased oncotic pressure in the peritubular capillaries will facilitate increased reabsorption of tubular fluid.
· Angiotensin II decreases medullary blood flow through the vasa recta. This decreases the washout of NaCl and urea in the kidney medullary space. Thus, higher concentrations of NaCl and urea in the medulla facilitate increased absorption of tubular fluid. Furthermore, increased reabsorption of fluid into the medulla will increase passive reabsorption of sodium along the thick ascending limb of the loop of Henle.
· Angiotensin II stimulates Na+/H+ exchangers located on the apical membranes (faces the tubular lumen) of cells in the proximal tubule and thick ascending limb of the loop of Henle in addition to Na+ channels in the collecting ducts. This will ultimately lead to increased sodium reabsorption
· Angiotensin II stimulates the hypertrophy of renal tubule cells, leading to further sodium reabsorption.
· In the adrenal cortex, it acts to cause the release of aldosterone. Aldosterone acts on the tubules (e.g., the distal convoluted tubules and the cortical collecting ducts) in the kidneys, causing them to reabsorb more sodium and water from the urine. This increases blood volume and, therefore, increases blood pressure. In exchange for the reabsorbing of sodium to blood, potassiumis secreted into the tubules, becomes part of urine and is excreted.
· Release of anti-diuretic hormone (ADH), also called vasopressin – ADH is made in the hypothalamus and released from the posterior pituitary gland. As its name suggests, it also exhibits vaso-constrictive properties, but its main course of action is to stimulate reabsorption of water in the kidneys. ADH also acts on the central nervous system to increase an individual's appetite for salt, and to stimulate the sensation of thirst.
These effects directly act in concert to increase blood pressure.
Local renin-angiotensin systems
Locally expressed renin-angiotensin systems have been found in a number of tissues, including the kidneys, adrenal glands, the heart, vasculature and nervous system, and have a variety of functions, including local cardiovascular regulation, in association or independently of the systemic renin-angiotensin system, as well as non-cardiovascular functions. Outside the kidneys, renin is predominantly picked up from the circulation but may be secreted locally in some tissues; its precursor prorenin is highly expressed in tissues and more than half of circulating prorenin is of extrarenal origin, but its physiological role besides serving as precursor to renin is still unclear.Outside the liver, angiotensinogen is picked up from the circulation or expressed locally in some tissues; with renin they form angiotensin I, and locally expressed angiotensin converting enzyme, chymase or other enzymes can transform it into angiotensin II. This process can be intracellular or interstitial.
In the adrenal glands, it is likely involved in the paracrine regulation of aldosterone secretion, in the heart and vasculature, it may be involved in remodeling or vascular tone, and in the brain where it is largely independent of the circulatory RAS, it may be involved in local blood pressure regulation. In addition, both the central and peripheral nervous systems can use angiotensin for sympathetic neurotransmision. Other places of expression include the reproductive system, the skin and digestive organs. Medications aimed at the systemic system may affect the expression of those local systems, beneficially or adversely.
|
|
Peculiarities of biochemical processes in kidney.
Kidney have a very high level of metabolic processes. They use about 10
% of all O2, which used in organism. During 24 hours through kidney
pass 700-
Metabolism of proteins also present in kidney in high level. Especially, glutamine deaminase is very active and a lot of free ammonia formed. In kidney take place the first reaction of creatin synthesis.
Kidney have plenty of different enzymes: LDG (1, 2, 3, 5), AsAT, AlAT. Specific for kidney is alanine amino peptidase, 3rd isoform.
Utilization of glucose in cortex and medulla is differ one from another. Dominative type of glycolysis in cortex is aerobic way and CO2 formed in result. In medulla dominative type is anaerobic and glucose converted to lactate.
Two sources contribute to the renal ammonia: blood ammonia (is about one-third of excreted ammonia), and ammonia formed in the kidney. The predominant source for ammonia production within the kidney is glutamine, the most abundant amino acid in plasma, but a small amount may originate from the metabolism of other amino acids such as asparagine, alanine, and histidine. Ammonia is secreted into the tubular lumen throughout the entire length of the nephron. Secretion occurs both during normal acid-base balance and in chronic acidosis.Metabolic acidosis is accompanied by an adaptive increase in renal ammonia production with a corresponding increase in urinary ammonium excretion.
Kidney cortex like liver appear to be unique in that it possess the enzymatic potential for both glucose synthesis from noncarbohydrate precursors (gluconeogenesis) and glucose degradation via the glycolytic pathway. Gluconeogenesis is important when the dietary supply of glucose does not satisfy the metabolic demands. Under these conditions, glucose is required by the central nervous system, the red blood cells, and possibly other tissues which cannot obtain all their energy requirements from fatty acids or ketone body oxidation. Also, gluconeogenesis may be important in the removal of excessive quantities of glucose precursors from the blood (lactate acid after severe exercise for example). Although the biosynthetic pathways are similar, there are several important differences in the factors which regulate gluconeogenesis in the two organs. 1) The liver utilizes predominately pyruvate, lactate and alanine. The kidney cortex utilizes pyruvate, lactate, citrate, α-ketoglutarate, glycine and glutamine. 2) Hydrogen ion activity has little effect upon hepatic gluconeogenesis, but it has marked effects upon renal gluconeogenesis. Thus, when intracellular fluid pH is reduced (metabolic acidosis, respiratory acidosis or potassium depletion), the rates of gluconeogenesis in slices of renal cortex are markedly increased. The ability of the kidney to convert certain organic acids to glucose, a neutral substance, is an example of a nonexcretory mechanism in the kidney for pH regulation.
Regulation of urine formation.
Na-uretic hormone (produced in heart) decrease reabsorbtion of Na+, and quantity of urine increased.
Aldosteron and some other hormones (vasopressin, renin, angiotensin II) increase Na-reabsorption and decrease quantity of urine.
Role of kidney in acid-base balance regulation.
The kidneys have two important roles in the maintaining of the acid-base balance: to reabsorb bicarbonate from and to excrete hydrogen ions into urine. 4500 mmol of bicarbonate are filtered into the primary filtrate of urine daily, but only 2 mmol of it are finally excreted. 70-80% of bicarbonate is reabsorbed in the first part of proximal tubule, 10-20% in the loop of Henle and 5-10% in the distal tubule and collecting ducts. (Jalanko & Holmberg 1998)
Carbonic anhydrase plays an important role in the reabsorption in the proximal tubule. Disturbance in the reabsorption of bicarbonate in the proximal tubule leads to metabolic acidosis, hyperchloremia and alkalotic urine. This disease is named as "type II renal tubular acidosis" (N25.8). (Jalanko & Holmberg 1998)
Renal tubules actively secrete hydrogen ions. Most of this takes place in the distal part of the nephron, but active transport of hydrogen ions occurs in the proximal tubule, too. The H-ATPase of the apical cell membrane secretes hydrogen ions into urine. For each hydrogen ion secreted, one bicarbonate molecule is transported to the interstitial fluid, from there it diffuses into the bloodstream. Fifty mmol of hydrogen ions are normally excreted daily. (Jalanko & Holmberg 1998)
If the hydrogen ions are not properly secreted into the collecting ducts, the result is metabolic acidosis, hypokalemia, hypocalcemia, nephrocalcinosis and an alkalotic urine. This disease is called "type I renal tubular acidosis" (N25.8). (Jalanko & Holmberg 1998)
The maximal
hydrogen ion gradient, against which the transport mechanism can secrete H+ ions, corresponds to a urine pH of
The source of the hydrogen ions secreted by the tubular cells is not completely certain. It is probably produced by dissociation of H2CO3. The acid-secreting cells contain carbonic anhydrase, which facilitates the rapid formation of H2CO3 from CO2 and water. The renal acid secretion is mainly regulated by the changes in the intracellular pCO2, potassium concentration, carbonic anhydrase activity and adrenocortical hormone concentration. (Ganong 1991)
Kidney have some mechanisms for maintaining acid-base balance. Na+ reabsorbtion and H+ secretion play very important role.
1. Primary urine has a lot of Na2HPO4 (in dissociated form). When Na+ reabsorbed, H+ secreted into urine and NaH2PO4 formed.
2. Formation of hydrocarbonates. Inside renal cells carboanhydrase forms from CO2 and H2O H2CO3, which dissociated to H+ and HCO3-. H+ excreted from cell into urine (antiport with Na+) and leaded with urine. Na+ connect with HCO3-, NaHCO3 formed and go to the blood, thereupon acidity decreased.
3. Formation of free ammonia. NH3 used for formation of NH4+ (H+ ion associted), and different acid metabolites excreted as ammonia salts.
The organs involved in regulation of external acid-base balance are the lungs are the kidneys.
The lungs are important for excretion of carbon dioxide (the respiratory acid) and there is a huge amount of this to be excreted: at least 12,000 to 13,000 mmols/day.
In contrast the kidneys are responsible for excretion of the fixed acids and this is also a critical role even though the amounts involved (70-100 mmols/day) are much smaller. The main reason for this renal importance is because there is no other way to excrete these acids and it should be appreciated that the amounts involved are still very large when compared to the plasma [H+] of only 40 nanomoles/litre.
There is a second extremely important role that the kidneys play in acid-base balance, namely the reabsorption of the filtered bicarbonate. Bicarbonate is the predominant extracellular buffer against the fixed acids and it important that its plasma concentration should be defended against renal loss.
In acid-base balance, the kidney is responsible for 2 major activities:
· Reabsorption of filtered bicarbonate: 4,000 to 5,000 mmol/day
· Excretion of the fixed acids (acid anion and associated H+): about 1 mmol/kg/day.
Both these processes involve secretion of H+ into the lumen by the renal tubule cells but only the second leads to excretion of H+ from the body.
The renal mechanisms involved in acid-base balance can be difficult to understand so as a simplification we will consider the processes occurring in the kidney as involving 2 aspects:
· Proximal tubular mechanism
· Distal tubular mechanism
The contributions of the proximal tubules to acid-base balance are:
· firstly, reabsorption of bicarbonate which is filtered at the glomerulus
· secondly, the production of ammonium
The next 2 sections explain these roles in more detail.
Daily filtered bicarbonate equals the product of the daily glomerular filtration rate (180 l/day) and the plasma bicarbonate concentration (24 mmol/l). This is 180 x 24 = 4320 mmols/day (or usually quoted as between 4000 to 5000 mmols/day).
About 85 to 90% of the filtered bicarbonate is reabsorbed in the proximal tubule and the rest is reabsorbed by theintercalated cells of the distal tubule and collecting ducts.
The reactions that occur are outlined in the diagram. Effectively, H+ and HCO3- are formed from CO2 and H2O in a reaction catalysed by carbonic anhydrase. The actual reaction involved is probably formation of H+ and OH- from water, then reaction of OH- with CO2 (catalysed by carbonic anhydrase) to produce HCO3-. Either way, the end result is the same.
The H+ leaves the proximal tubule cell and enters the PCT lumen by 2 mechanisms:
· Via a Na+-H+ antiporter (major route)
· Via H+-ATPase (proton pump)
Filtered HCO3- cannot cross the apical membrane of the PCT cell. Instead it combines with the secreted H+ (under the influence of brush border carbonic anhydrase) to produce CO2 and H2O. The CO2 is lipid soluble and easily crosses into the cytoplasm of the PCT cell. In the cell, it combines with OH- to produce bicarbonate. The HCO3-crosses the basolateral membrane via a Na+-HCO3- symporter. This symporter is electrogenic as it transfers three HCO3- for every one Na+. In comparison, the Na+-H+ antiporter in the apical membrane is not electrogenic because an equal amount of charge is transferred in both directions.
The basolateral membrane also has an active Na+-K+ ATPase (sodium pump) which transports 3 Na+ out per 2 K+ in. This pump is electrogenic in a direction opposite to that of the Na+-HCO3- symporter. Also the sodium pump keeps intracellular Na+ low which sets up the Na+ concentration gradient required for the H+-Na+ antiport at the apical membrane. The H+-Na+ antiport is an example of secondary active transport.
The net effect is the reabsorption of one molecule of HCO3 and one molecule of Na+ from the tubular lumen into the blood stream for each molecule of H+ secreted. This mechanism does not lead to the net excretion of any H+ from the body as the H+ is consumed in the reaction with the filtered bicarbonate in the tubular lumen.
[Note: The differences in functional properties of the apical membrane from that of the basolateral membranes should be noted. This difference is maintained by the tight junctions which link adjacent proximal tubule cells. These tight junctions have two extremely important functions:
Gate function: They limit access of luminal solutes to the intercellular space. This resistance can be altered and this paracellular pathway can be more open under some circumstances (ie the ‘gate’ can be opened a little).
Fence function: The junctions maintain different distributions of some of the integral membrane proteins. For example they act as a ‘fence’ to keep the Na+-H+ antiporter limited to the apical membrane, and keep the Na+-K+ ATPase limited to the basolateral membrane. The different distribution of such proteins is absolutely essential for cell function.]
The 4 major factors which control bicarbonate reabsorption are:
· Luminal HCO3- concentration
· Luminal flow rate
· Arterial pCO2
· Angiotensin II (via decrease in cyclic AMP)
An increase in any of these four factors causes an increase in bicarbonate reabsorption. Parathyroid hormone also has an effect: an increase in hormone level increases cAMP and decreases bicarbonate reabsorption.
The mechanism for H+ secretion in the proximal tubule is described as a high capacity, low gradient system:
The high capacity refers to the large amount (4000 to 5000 mmols) of H+ that is secreted per day. (The actual amount of H+ secretion is 85% of the filtered load of HCO3-).
The low gradient refers to the low pH gradient as tubular pH can be decreased from 7.4 down to 6.7-7.0 only.
Though no net excretion of H+ from the body occurs, this proximal mechanism is extremely important in acid-base balance. Loss of bicarbonate is equivalent to an acidifying effect and the potential amounts of bicarbonate lost if this mechanism fails are very large.
Ammonium (NH4) is produced predominantly within the proximal tubular cells. The major source is from glutamine which enters the cell from the peritubular capillaries (80%) and the filtrate (20%). Ammonium is produced from glutamine by the action of the enzyme glutaminase. Further ammonium is produced when the glutamate is metabolised to produce alpha-ketoglutarate. This molecule contains 2 negatively-charged carboxylate groups so further metabolism of it in the cell results in the production of 2 HCO3- anions. This occurs if it is oxidised to CO2 or if it is metabolised to glucose.
The pKa for ammonium is so high that both at extracellular and at intracellular pH, it is present entirely in the acid form NH4+. The previous idea that lipid soluble NH3 is produced in the tubular cell, diffuses into the tubular fluid where it is converted to water soluble NH4+ which is now trapped in the tubule fluid is incorrect.
The subsequent situation with ammonium is complex. Most of the ammonium is involved in cycling within the medulla. About 75% of the proximally produced ammonium is removed from the tubular fluid in the medulla so that the amount of ammonium entering the distal tubule is small. The thick ascending limb of the loop of Henle is the important segment for removing ammonium. Some of the interstitial ammonium returns to the late proximal tubule and enters the medulla again (ie recycling occurs).
An overview of the situation so far is that:
· The ammonium level in the DCT fluid is low because of removal in the loop of Henle
· Ammonium levels in the medullary interstitium are high (and are kept high by the recycling process via the thick ascending limb and the late PCT)
· Tubule fluid entering the medullary collecting duct will have a low pH if there is an acid load to be excreted (and the phosphate buffer has been titrated down.
If H+ secretion continues into the medullary collecting duct this would reduce the pH of the luminal fluid further. A low pH greatly augments transfer of ammonium from the medullary interstitium into the luminal fluid as it passes through the medulla. The lower the urine pH, the higher the ammonium excretion and this ammonium excretion is augmented further if an acidosis is present. This augmentation with acidosis is 'regulatory' as the increased ammonium excretion by the kidney tends to increase extracellular pH towards normal.
If the ammonium returns to the blood stream it is metabolised in the liver to urea (Krebs-Henseleit cycle) with net production of one hydrogen ion per ammonium molecule.
Regulation of Renal H+ Excretion
The discussion above has described the mechanisms involved in renal acid excretion and mentioned some factors which regulate acid excretion.
The major factors which regulate renal bicarbonate reabsorption and acid excretion are:
1. Extracellular volume
Volume depletion is associated with Na+ retention and this also enhances HCO3 reabsorption. Conversely, ECF volume expansion results in renal Na+ excretion and secondary decrease in HCO3 reabsorption.
2. Arterial pCO2
An increase in arterial pCO2 results in increased renal H+ secretion and increased bicarbonate reabsorption. The converse also applies. Hypercapnia results in an intracellular acidosis and this results in enhanced H+ secretion. The cellular processes involved have not been clearly delineated. This renal bicarbonate retention is the renal compensation for a chronic respiratory acidosis.
3. Potassium & Chloride Deficiency
Potassium has a role in bicarbonate reabsorption. Low intracellular K+ levels result in increased HCO3 reabsorption in the kidney. Chloride deficiency is extremely important in the maintenance of a metabolic alkalosis because it prevents excretion of the excess HCO3 (ie now the bicarbonate instead of chloride is reabsorbed with Na+ to maintain electroneutrality).
4. Aldosterone & cortisol (hydrocortisone)
Aldosterone at normal levels has no role in renal regulation of acid-base balance. Aldosterone delpetion or excess does have indirect effects. High aldosterone levels result in increased Na+ reabsorption and increased urinary excretion of H+ and K+ resulting in a metabolic alkalosis. Conversely, it might be thought that hypoaldosteronism would be associated with a metabolic acidosis but this is very uncommon but may occur if there is coexistent significant interstitial renal disease.
5. Phosphate Excretion
Phosphate is the major component of titratable acidity. The amount of phosphate present in the distal tubule does not vary greatly. Consequently, changes in phosphate excretion do not have a significant regulatory role in response to an acid load.
6. Reduction in GFR
It has recently been established that a reduction in GFR is a very important mechanism responsible for the maintenance of a metabolic alkalosis. The filtered load of bicarbonate is reduced proportionately with a reduction in GFR.
7. Ammonium
The kidney responds to an acid load by increasing tubular production and urinary excretion of NH4+. The mechanism involves an acidosis-stimulated enhancement of glutamine utilisation by the kidney resulting in increased production of NH4+ and HCO3- by the tubule cells. This is very important in increasing renal acid excretion during a chronic metabolic acidosis. There is a lag period: the increase in ammonium excretion takes several days to reach its maximum following an acute acid load. Ammonium excretion can increase up to about 300 mmol/day in a chronic metabolic acidosis so this is important in renal acid-base regulation in this situation. Ammonium excretion increases with decreases in urine pH and this relationship is markedly enhanced with acidosis.
![]()
Role of kidney in water balance regulation.
Excessive entrance of water leads to dilution of extracellular fluid. Decreasing of osmolality inhibits secretion of antidiuretic hormone. Walls of collective tubules stay non-penetrated to water and dilutive urine formed.
If volume of blood circulation increases, circulation in kidney increases also and hyperosmotic medium of kidney medulla removed. Some substances in these conditions return into blood. So, excess of water carried with urine and a lot of soluble substances are reabsorbed into blood. After water loading stopped, hyperosmolality in kidney medulla returns for previous stage during some days.
Physical and chemical characteristics of urine.
Urine amount (diures) in healthy people is 1000-2000 ml per day. Day-time diures is in 3-4 times more than night-time.
Normal colour of urine is yellow (like hay or amber), what is due to presence of urochrom (derivate of urobilin or urobilinogen). Some another colour substances are uroerythrin (derivate of melanine), uroporphyrines, rybophlavine and other. Colour depends from urine concentration.
Urine varies in appearance, depending principally upon a body's level of hydration, as well as other factors. Normal urine is a transparent solution ranging from colorless to amber but is usually a pale yellow. In the urine of a healthy individual the color comes primarily from the presence of urobilin. Urobilin in turn is a final waste product resulting from the breakdown of heme from hemoglobin during the destruction of aging blood cells.
Colorless urine indicates over-hydration, generally preferable to dehydration (though it can remove essential salts from the body). Colorless urine in drug tests can suggest an attempt to avoid detection of illicit drugs in the bloodstream through over-hydration.
· Dark yellow urine is often indicative of dehydration.
· Yellowing/light orange may be caused by removal of excess B vitamins from the bloodstream.
· Certain medications such as rifampin and phenazopyridine can cause orange urine.
· Bloody urine is termed hematuria, a symptom of a wide variety of medical conditions
· Dark orange to brown urine can be a symptom of jaundice, rhabdomyolysis, or Gilbert's syndrome.
· Black or dark-colored urine is referred to as melanuria and may be caused by a melanoma.
· Pinkish urine can result from the consumption of beets.
· Greenish urine can result from the consumption of asparagus.
· Reddish or brown urine may be caused by porphyria (not to be confused with the harmless, temporary pink or reddish tint caused by beeturia).
· Blue urine can be caused by the ingestion of methylene blue, e.g. in medications
· Blue urine stains can be caused by blue diaper syndrome
Urine is transparent. This characteristic depends from amount of different salts (oxalates, urates, phosphates), amount of present epitelium cells and leucocytes.
Density of urine depends from concentration of soluble substances.
Borders of variation are from 1002 to 1035 g/l. Near 60-
The pH of urine can vary between 4.6 and 8, with neutral (7) being norm. In persons with hyperuricosuria, acidic urine can contribute to the formation of stones of uric acid in the kidneys, ureters, or bladder. Urine pH can be monitored by a physician or at home.
A diet high in citrus, vegetables, or dairy can increase urine pH (more basic). Some drugs also can increase urine pH, including acetazolamide, potassium citrate, and sodium bicarbonate.
A diet high in meat can decrease urine pH (more acidic). Cranberries, popularly thought to decrease the pH of urine, have actually been shown not to acidify urine. Drugs that can decrease urine pH include ammonium chloride, chlorothiazide diuretics, and methenamine mandelate.[14][15]
In normal conditions urine has acid or weak acid reaction (pH=5,3-6,8). This depends from presence of NaH2PO4 and KH2PO4.
Fresh urine has a specific smell, which is due to presence of flying acids. But a lot of microorganisms, which are present in urine, split urea and free ammonia formed.
Exhaustive detailed description of the composition of human urine can be found in NASA Contractor Report No. NASA CR-1802, D. F. Putnam, July 1971. That report provided detailed chemical analyses for inorganic and organic constituents, methods of analysis, chemical and physical properties and its behavior during concentrative processes such as evaporation, distillation and other physiochemical operations. Urine is an aqueous solution of greater than 95% water, with the remaining constituents, in order of decreasing concentration urea 9.3 g/L, chloride 1.87 g/L, sodium 1.17 g/L, potassium 0.750 g/L, creatinine 0.670 g/L and other dissolved ions, inorganic and organic compounds.
Urine is sterile until it reaches the urethra, where epithelial cells lining the urethra are colonized by facultatively anaerobic Gram negative rods and cocci. Subsequent to elimination from the body, urine can acquire strong odors due to bacterial action, and in particular the release of ammonia from the breakdown of urea.
Some diseases alter the quantity and consistency of urine, such as diabetes introducing sugar. Consuming beets can result in beeturia (pink/red urine containing betanin) for some 10–14% of the popu Urine is a liquid produced by the kidneys to remove waste products from the bloodstream. Human urine is yellowish in color and variable in chemical composition, but here is a list of its primary components.
Human urine consists primarily of water, with organic solutes including urea, creatinine, uric acid, and trace amounts of enzymes, carbohydrates, hormones, fatty acids, pigments, and mucins, and inorganic ions such as sodium (Na+), potassium (K+), chloride (Cl-), magnesium (Mg2+), calcium (Ca2+), ammonium (NH4+), sulfates (SO42-), and phosphates (e.g., PO43-). A representative chemical composition would be:
water 95%
urea 9.3 g/l
chloride 1.87 g/l
sodium 1.17 g/l
potassium 0.750 g/l
creatinine 0.670 g/l
Organic compounds of urine.
Proteins. Healthy people excretes 30 mg of proteins per day. As a rule these are low weight proteins.
Urea. This is main part of organic compounds in urine. Urea nitrogen is about
80-90 % of all urine nitrogen. 20-
Uric acid. Approximately 0,6-1,0 g of uric acid is excreted per day in form of different salts (urates), mainly in form of sodium salt. Its amount depends from food.
Creatinin and
creatin. Near 1-
Amino acids. Per day healthy person excretes 2-
Couple substances. Hypuric acid (benzoyl glycine) is excreted in amount 0,6-
Indican (potassium
salt of indoxylsulfuric acid). Per day excrition of indican
is about 10-
Organic acids. Formic, acetic, butyric, β-oxybutyric, acetoacetic and some other organic acids are present in urine in a little amount.
Vitamines. Almost all vitamines can be excreted via kidney, especially, water-soluble. Approximately 20-30 mg of vit C, 0.1-0.3 mg of vit B1, 0.5-0.8 mg of vit B2 and some products of vitamine’s metabolism. These data can be used for evaluating of supplying our organism by vitamines.
Hormones. Hormones and their derivates are always present in urine. Their amount
depends from functional state of endocrinal glands and liver. There is a very
wide used test – determination of 17-ketosteroids in urine. For healthy man this
index is 15-
Urobilin. Present in a little amount, gives to urine yellow colour.
Bilirubin. In normal conditions present in so little amount that cannot be found by routine methods of investigations.
Glucose. In normal conditions present in so little amount that cannot be found by routine methods of investigations.
Galactose. Present in the newborn’s urine, when digestion of milk or transformation of glalactose into glucose in the liver are violated.
Fructose. It is present in urine very seldom, after eating a lot of fruits, berries and honey. In all other cases it indicates about liver’s disorders, diabetes mellitus.
Pentoses. Pentoses are excreted after eating a lot of fruits, fruit juices, in case of diabetes mellitus and steroid diabetes, some intoxications.
Ketone bodies. In normal conditions urine contains 20-50 mg of ketone bodies and this amount cannot be found by routine methods of clinical investigations.
Porphyrines. Urine of healthy people contains a few I type porphyrines (up to 300 mkg per day).
Inorganic compounds of urine.
Urine of healthy people contains 15-
NaCl. Per day near 8-
Potassium. Twenty-four hours urine contains 2-
Different drugs can change excretion of Na and K. For example, salicylates and cortikosteroids keep Na and amplify excretion of K.
Calcium. Twenty-four hours urine contains 0.1-
Magnesium. Content of magnesium in urine is 0.03-
Iron. Amount of iron in urine is about 1 mg per day.
Phosphorus. In urine are present one-substituted phosphates of potassium and sodium. Their amount depends from blood pH. In case of acidosis two-substituted phosphates (Na2HPO4) react with H+ and one-substituted phosphates (NaH2PO4) formed. In case of alkalosis one-substituted phosphates react with bases and two-substituted phosphates formed. So, in both cases amount of phosphates in urine increases.
Sulfur. Amount of sulfur in twenty-four hours urine is 2-
Ammonia. Ammonia is excreted in ammonium sulfates and couple substances. Ammonium salts make up 3-6 % of all nitrogen in urine. Their amount depends from character of food and blood pH.
Urine analysis infers valuable information in a variety of ailments. Physical characteristics of urine have been used as diagnostic and prognostic tool from the time immemorial by the health physicians. We know that the major functions of kidneys are:
· Removal of water not needed by the body fluids, the amount depending on the balance between glomerular filtrate and he degree of tubular reabsorption;
· The excretion of certain substances normally present in the plasma when their concentration rises above a certain level;
· The selective reabsorption of substances such as glucose which are of value to the body;
· The excretion of useless substances; and
· Regulation of acid base balance.
Disordered renal function may lead to a change in the volume of the urine excreted per day along with remarkable changes in its physical and chemical properties and microscopic contents. Urine analysis is the very first investigation of diagnostic importance not only in renal disorders but also in other diseases like diabetes, liver disease, jaundice etc. In diagnostic pathology the extent of abnormalities could only be understood in comparison with the reference values obtained from similar investigations in normal individuals. Hence, it is important to have an understanding of normal parameters of physical and chemical characteristics of urine.
Characteristics of normal urine:
· Quantity: The quantity averages 1500 to 2000 ml in an adult man daily. It may vary with the amount of fluid taken. In fact it is linked with the protein metabolism; higher is the protein intake higher will be the urinary output since the urea produced from the protein needs to be flushed out from the body. Higher is the urea production in the body, the higher is the volume of urine to excrete it.
· Color: The color should be clear pale amber without any deposits. However, a light flocculent cloud of mucus may sometimes be seen floating in the normal urine.
· Specific gravity: It varies from 1.010 to 1.025. Specific gravity is determined with urinometer.
· Odor: The odor is aromatic.
· Reaction: The reaction of normal urine is slightly acidic with an average pH of 6.0.
Composition of normal urine: Urine is mainly composed of
water, urea and sodium chloride. I an adult taking about
· Water: Near about 96%
· Solids: About 4% (urea 2% and other metabolic products 2%. Other metabolic products include: uric acid, creatinine, electrolytes or salts such as sodium chloride, potassium chloride and bicarbonate).
§
Urea is one of the end products of protein metabolism. It is
prepared from the deaminated amino-acid in the liver and reach
the kidneys through blood circulation (The normal blood urea level is 20-40
mg/dl). About
§
Uric Acid: The normal level of uric acid in blood is 2 to 6 mg/dl
and about 1.5 to
§ Creatinine: Creatinine is the metabolic waste of creatin in muscle. Purine bodies, oxalates, phosphates, sulphates and urates are the other metabolic products.
§ Electrolytes or salts such as sodium chloride and potassium chloride are also excreted in the urine to maintain the normal level in blood. These are the salts which are the part of our daily diet and are always taken in excess and need to be excreted to maintain normal physiological balance.
Indeces of renal functions disorders.
Proteinuria. Proteinuria can be kidney and extralidney origin. Kidney proteinuria is due to damages of nephrons, when blood plasma proteins can pass through glomerular membranes. In this case albumins and globulins are present in urine. Extra kidney proteinuria is due to damages of urinary tracts and prostate.
Hematuria. This pathological component is due to damages of kidney or urinary tracts. In most cases nephrolytiasis is accompanied by hematuria. Some times hematuria is indicator of traumatical kidney damage.
Glucosuria. In most cases glucosuria is a symptom of diabetes mellitus, when level of glucose in blood more than kidney threshold (8-10 mmol/L). But sometimes glucose can be present in urine even its level in blood is normal. This is so called “renal glucosuria” which is due to disorders of glucose reabsorbtion in tubules.
Pyuria. In normal urine leucocytes are present in a very little amount. Due to different inflammatory processes of urinary tracts, urine bladder, prostate, also due to nephrolytiasis amount of leucocytes in urine increases and this situation is called “pyuria”.
Creatin. Decreasing of creatin in urine is index of kidney insufficiency.
Proteinuria (PU)
• Means pathological presence of protein in definitive urine (i.e. > 0.15 g/24 hr.)
• Is common sign of glomerular impairment:
• Mechanical: non-selective proteinuria
• Charge loss: selective PU - albuminuria
• Tubular proteinuria: the presence of proteins normally filtered, which are not
resorbed in tubules
• Extrarenal causes: myoglobine, Hb e
Proteinuria (from protein and urine) means the presence of an excess of serum proteins in the urine. The excess protein in the urine often causes the urine to become foamy, although foamy urine may also be caused by bilirubin in the urine (bilirubinuria), retrograde ejaculation, pneumaturia (air bubbles in the urine) due to a fistula, or drugs such as pyridium.
Proteinuria may be a sign of renal (kidney) damage. Since serum proteins are readily reabsorbed from urine, the presence of excess protein indicates either an insufficiency of absorption or impaired filtration. Diabetics may suffer from damaged nephrons and develop proteinuria. The most common cause of proteinuria is diabetes, and in any person with proteinuria and diabetes, the etiology of the underlying proteinuria should be separated into two categories: diabetic proteinuria versus the field.
With severe proteinuria, general hypoproteinemia can develop which results in diminished oncotic pressure. Symptoms of diminished oncotic pressure may include ascites, edema andhydrothorax.
Conditions with proteinuria as a sign
Proteinuria may be a feature of the following conditions:
· Nephrotic syndromes (i.e. intrinsic renal failure)
· Toxic lesions of kidneys
· Amyloidosis
· Collagen vascular diseases (e.g. systemic lupus erythematosus)
· Dehydration
· Glomerular diseases, such as membranous glomerulonephritis, focal segmental glomerulonephritis, minimal change disease (lipoid nephrosis)
· Strenuous exercise
· Stress
· Benign orthostatic (postural) proteinuria
· Focal segmental glomerulosclerosis (FSGS)
· IgA nephropathy (i.e. Berger's disease)
· Membranoproliferative glomerulonephritis
· Diabetes mellitus (diabetic nephropathy)
· Drugs (e.g. NSAIDs, nicotine, penicillamine, lithium carbonate, gold and other heavy metals, ACE inhibitors, antibiotics, or opiates (especially heroin)[16]
· Infections (e.g. HIV, syphilis, hepatitis, poststreptococcal infection, urinary schistosomiasis)
· Hypertensive nephrosclerosis
· Organ rejection: Kidney transplant patients may have gamma-globulins in their urine if the kidneys start to reject.[17]
· Familial Mediterranean fever
· Systemic lupus erythematosus
· Rheumatoid arthritis
· Glycogen storage disease type 1
· A urinary tract infection which has spread to the kidney(s)
This list is incomplete; you can help by expanding it.
Conditions with proteinuria consisting mainly of Bence-Jones proteins as a sign
· Waldenstrom's macroglobulinemia
· Chronic lymphocytic leukemia
· Malignancies (e.g., lymphoma, other cancers)
There are three main mechanisms to cause proteinuria:
· Due to disease in glomerulus
· Because of increased quantity of proteins in serum (overflow proteinuria)
· Due to low reabsorption at proximal tubule (Fanconi syndrome)
Proteinuria can also be caused by certain biological
agents, such as bevacizumab (Avastin) used in cancer treatment, or
by excessive fluid intake (drinking in excess of
Glycosuria or glucosuria is the excretion of glucose into the urine. Ordinarily, urine contains no glucose because the kidneys are able to reclaim all of the filtered glucose back into the bloodstream. Glycosuria is nearly always caused by elevated blood glucose levels, most commonly due to untreated diabetes mellitus. Rarely, glycosuria is due to an intrinsic problem with glucose reabsorption within the kidneys themselves, a condition termed renal glycosuria. Glycosuria leads to excessive water loss into the urine with resultant dehydration, a process called osmotic diuresis.
Glycosuria refers to sugar in the urine. Less than 0.1% of glucose normally filtered by the glomeruli appears in the urine, and less than 130 mg should appear in the urine over a 24-hour period. Glucose is present in glomerular filtrate but is reabsorbed by the kidney's proximal tubule. If the blood glucose level exceeds the capacity of the tubules to reabsorb all the glucose present in the glomerular filtrate, the renal threshold is reached and glucose spills into the urine. A finding of glycosuria indicates that the person is hyperglycemic or has a lowered renal threshold for glucose. The renal threshold for glucose is approximately 160 to 190mg/dl of blood; glucose does not appear in the urine until the blood glucose rises above this level.
Occasionally, glycosuria may be a normal finding, such as after eating a heavy meal or during times of emotional stress. Some individuals have a benign condition in which they have a lower than usual renal threshold for glucose, but have normal blood glucose levels. In pregnancy, the renal threshold for glucose may be lowered so that small amounts of glycosuria may be present. Patients on hyperalimentation may have glycosuria if the carbohydrate solution is being infused faster than the pancreas can produce insulin. The most common reason for glycosuria is diabetes mellitus. Urine glucose tests are used to screen for diabetes, to confirm a diagnosis of diabetes, or to monitor diabetic control.
Renal glycosuria, also known as renal glucosuria, is a rare condition in which the simple sugar glucose is excreted in the urine despite normal or low blood glucose levels. With normal kidney (renal) function, glucose is excreted in the urine only when there are abnormally elevated levels of glucose in the blood. However, in those with renal glycosuria, glucose is abnormally elevated in the urine due to improper functioning of the renal tubules, which are primary components of nephrons, the filtering units of the kidneys.
Hematuria, or haematuria, is the presence of red blood cells (erythrocytes) in the urine. It may be idiopathic and/or benign, or it can be a sign that there is a kidney stone or a tumor in the urinary tract (kidneys, ureters, urinary bladder, prostate, and urethra), ranging from trivial to lethal. If white blood cells are found in addition to red blood cells, then it is a signal of urinary tract infection.
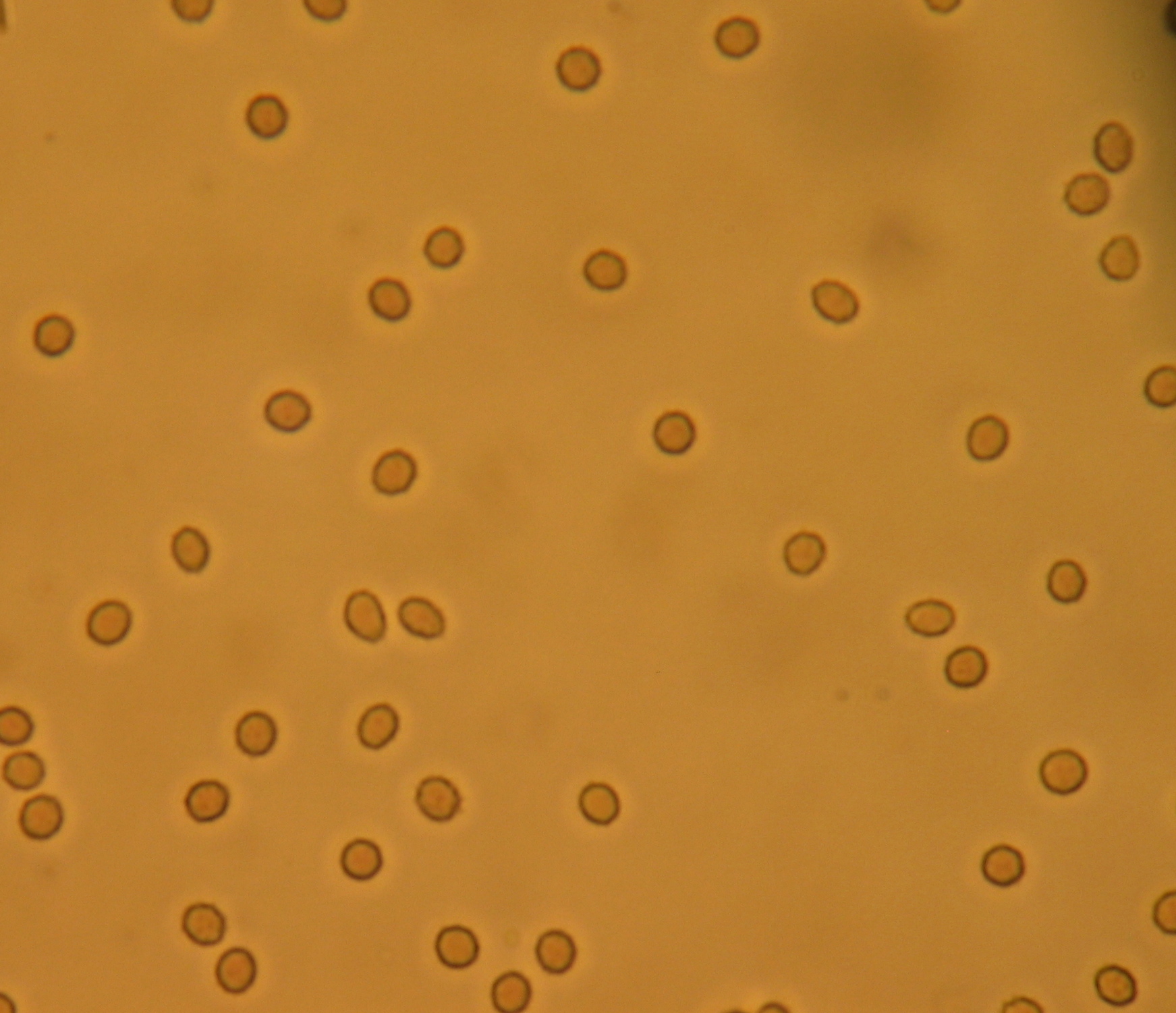
Microscopic hematuria
Occasionally "hemoglobinuria" is used synonymously, although more precisely it refers only to hemoglobin in the urine.
Red discoloration of the urine can have various causes:
- Red blood cells
- Microscopic hematuria (small amounts of blood, can be seen only on urinalysis or light microscopy)
- Macroscopic hematuria (or "frank" or "gross") hematuria
- Hemoglobin (only the red pigment, not the red blood cells)
- Other pigments
- Myoglobin in myoglobinuria
- Porphyrins in porphyria
- Betanin, after eating beets
- Drugs such as Rifampicin and Phenazopyridine
Diagnosis

Acute hematuria due to trauma.
Often, the diagnosis is made on the basis of the medical history and some blood tests—especially in young people in whom the risk of malignancy is negligible and the symptoms are generally self-limiting.
Ultrasound investigation of the renal tract is often used to distinguish between various sources of bleeding. X-rays can be used to identify kidney stones, although CT scanning is more precise.
In older patients, cystoscopy with biopsy of suspected lesions is often employed to investigate for bladder cancer.
If combined with pain, it may be loin pain hematuria syndrome.
The dipstick method of chemical analysis of urine
Pathological components of urine, which are occur due to different metabolic disorders in organism.
Ketonuria. Due to some diseases and pathological conditions (diabetes mellitus, starvation, severe heart weakness, when amount of fat in the food more than amount of carbohydrates) level of ketone bodies increases in a big amount (up to 20-50 g per day). This is the index of deep metabolic disorders, especially in carbohydrates metabolism.
Bilirubinuria. It occurs in case of hepatic parenhimatous inflammatory processes or in case of obstruction of gall bladder ductus. Urine has a colour as dark bear. After some times it stays yellow-green (bilirubin oxidized to biliverdin).
Urobilinuria. Increasing of amount of urobilin is due to haemolytic or parenhimatous hepatitis, when decomposition of mesobilinogen in liver is depressed.
Creatinuria. Amount of creatin in urine increases due to different pathological processes in muscles like myopathy and myodistrophy, starvation, hypovitaminosis E, radiation sickness, hyperthyreosis. Also this is present in small children and in women after delivery.
Indicanuria. Increasing of indican’s level in urine is due to intensification of decay proteins in the intestines, weaken of intestine peristaltic (atony, constipation), and chronic diseases, which are accompanied by intensive decomposition of proteins (tuberculosis, for example).
Phenylketonuria. Innate deficiency of phenylalanine hydroxilase in liver makes transformation of phenylalalnine in tyrosine impossible.