BIOCHEMISTRY AND PATHOBIOCHEMISTRY OF
BLOOD. RESPIRATORY FUNCTION OF ERYTHROCYTES. PATHOLOGICAL FORMS OF HEMOGLOBIN.
Blood is a liquid tissue. Suspended in the watery plasma are seven
types of cells and cell fragments.
Biological functions
of the blood
The blood is the most specialized fluid tissue which
circulates in vascular system and together with lymph and intercellular space
compounds an internal environment of an organism.
The blood executes
such functions:
1. Transport of gases – oxygen from lungs is
carried to tissues and carbon dioxide from tissues to lungs.
2. Transport of nutrients to all cells of
organism (glucose, amino acids, fatty acids, vitamins, ketone bodies, trace
substances and others). Substances such as urea, uric acid, bilirubin and
creatinine are taken away from the different organs for ultimate excretion.
3. Regulatory or hormonal function –
hormones are secreted in to blood and they are transported by blood to their
target cells.
4. Thermoregulation function - an exchange of
heat between tissues and blood.
5. Osmotic function- sustains osmotic pressure in
vessels.
6. Protective function- by the phagocytic action
of leucocytes and by the actions of antibodies, the blood provides the most
important defense mechanism.
7. Detoxification function - neutralization of
toxic substances which is connected with their decomposition by the help of
blood enzymes.
Blood performs two major
functions:
· transport through the body of
o oxygen and carbon dioxide
o food molecules (glucose, lipids, amino
acids)
o ions (e.g., Na+, Ca2+,
HCO3−)
o wastes (e.g., urea)
o hormones
o heat
· defense of
the body against infections and other foreign materials. All the WBCs participate in these defenses.
The formation of blood cells (cell types and
acronyms are defined below)
All the various types of blood cells

Are produced in the bone marrow (some
1011 of them each day in an adult human!). Arise from a single type of cell
called a hematopoietic stem cell — an "adult" multipotent stem cell.
These stem cells
• are very rare (only about one in 10,000 bone marrow
cells);
• are attached (probably by adherens junctions) to
osteoblasts lining the inner surface of bone cavities;
• express a cell-surface protein designated CD34;
•produce, by mitosis, two kinds of progeny:
o more stem cells (A mouse that has had all its blood
stem cells killed by a lethal dose of radiation can be saved by the injection
of a single living stem cell!).
o cells that
begin to differentiate along the paths leading to the various kinds of blood
cells.
Which path is taken is regulated by
• the need for more of that type of blood cell which
is, in turn, controlled by appropriate cytokines and/or hormones.
Examples:
Interleukin-7 (IL-7) is the major cytokine in
stimulating bone marrow stem cells to start down the path leading to the
various lymphocytes (mostly B cells andT
cells ).
Erythropoietin (EPO), produced by the kidneys,
enhances the production of red blood cells (RBCs).
Thrombopoietin (TPO), assisted by Interleukin-11
(IL-11), stimulates the production of megakaryocytes. Their fragmentation
produces platelets.
Granulocyte-macrophage colony-stimulating factor
(GM-CSF), as its name suggests, sends cells down the path leading to both those
cell types. In due course, one path or the other is taken.
Under the influence of granulocyte colony-stimulating
factor (G-CSF), they differentiate into neutrophils.
Further stimulated by interleukin-5 (IL-5) they
develop into eosinophils.
Interleukin-3 (IL-3) participates in the
differentiation of most of the white blood cells but plays a particularly
prominent role in the formation of basophils(responsible for some allergies).
Stimulated by macrophage colony-stimulating factor
(M-CSF) the granulocyte/macrophage progenitor cells differentiate into
monocytes, macrophages, and dendritic cells (DCs).
Biological chemistry of blood cells
Two types of blood cells can be distinguished - white and red blood
cells. White blood cells are called leucocytes. Their quantity in adult is 4-9 x 109/L. Red blood cells are called
erythrocytes. Their quantity in peripheral blood is 4,5-5 x 1012/L. Besides
that, there are also thrombocytes or platelets in blood.
White Blood Cells (leukocytes)
Leucocytes (white blood cells) protect an
organism from microorganisms, viruses and foreign substances, that provides the
immune status of an organism.
• are much less numerous than red
(the ratio between the two is around 1:700),
• have nuclei,
•
participate in protecting the body from infection,
• consist of lymphocytes and
monocytes with relatively clear cytoplasm, and three types of granulocytes,
whose cytoplasm is filled with granules.
Leucocytes are divided into two
groups: Granulocytes and agranulocytes. Granulocytes consist of neutrophils,
eosinophils and basophils. Agranulocytes consist of monocytes and lymphocytes.
Neutrophils
Neutrophils comprise of 60-70 % from
all leucocytes. Their main function is to protect organisms from microorganisms
and viruses. Neutrophils have segmented nucleus, endoplasmic reticulum
(underdeveloped) which does not contain ribosomes, insufficient amount of
mitochondria, well-developed Golgi apparatus and hundreds of different vesicles
which contain peroxidases and hydrolases. Optimum condition for their activity
is acidic pH. There are also small vesicles which contain alkaline
phosphatases, lysozymes, lactopherins and proteins of cationic origin.
Glucose is the main source of energy
for neutrophils. It is directly utilized or converted into glycogen. 90 % of
energy is formed in glycolysis, a small amount of glucose is converted in
pentosophosphate pathway. Activation of proteolysis during phagocytosis as well
as reduction of phosphatidic acid and phosphoglycerols are also observed. The
englobement is accompanied by intensifying of a glycolysis and pentosophosphate
pathway. But especially intensity of absorption of oxygen for neutrophils -
so-called flashout of respiration grows. Absorbed oxygen is spent for formation
of its fissile forms that is carried out with participation enzymes:
1. NADP*Н -OXYDASE catalyzes
formation of super oxide anion
2. An enzyme NADH- OXYDASE is responsible
for formation of hydrogen peroxide
3. Мyeloperoxydase catalyzes
formation of hypochloric acid from chloride and hydrogen peroxide
Neutrophils are motile phagocyte
cells that play a key role in acute inflammation. When bacteria enter tissues,
a number of phenomena occur that are collectively known as acute inflammatory
response. When neutrophils and other phagocyte cells engulf bacteria, they
exhibit a rapid increase in oxygen consumption known as the respiratory burst.
This phenomenon reflects the rapid utilization of oxygen (following a lag of
15-60 seconds) and production from it of large amounts of reactive derivates,
such as O2-, H2O2, OH. and OCl-
(hypochlorite ion). Some of these products are potent microbicidal
agents. The electron transport chain system responsible for the respiratory
burst contains several components, including a flavoprotein
NADPH:O2-oxidoreductase (often called NADPH-oxidase) and a b-type cytochrome.

The most abundant of the WBCs. This
photomicrograph shows a single neutrophil surrounded by red blood cells.
Neutrophils squeeze through the capillary
walls and into infected tissue where they kill the invaders (e.g., bacteria)
and then engulf the remnants by phagocytosis.
This is a never-ending task, even in
healthy people: Our throat, nasal passages, and colon harbor vast numbers of
bacteria. Most of these are commensals, and do us no harm. But that is because
neutrophils keep them in check.
However,
• heavy doses of radiation
• chemotherapy
• and many other forms of
stress
can reduce the numbers of neutrophils so
that formerly harmless bacteria begin to proliferate. The resulting
opportunistic infection can be life-threatening.
http://www.youtube.com/watch?v=EpC6G_DGqkI&feature=related
Some important enzymes and proteins of
neutrophilis.
Myeloperoxidase (MPO). Catalyzed following
reaction:
H2O2 + X-(halide) + H+® HOX + H2O (where
X- = Cl-, Br-, I- or SCN-; HOX=hypochlorous acid)
HOCl, the active ingredient of household
liquid bleach, is a powerful oxidant and is highly microbicidial. When applied
to normal tissues, its potential for causing damage is diminished because it
reacts with primary or secondary amines present in neutrophils and tissues to
produce various nitrogen-chlorine (N-Cl) derivates; these chloramines are also
oxidants, although less powerful than HOCl, and act as microbicidial agents
(eg, in sterilizing wounds) without causing tissue damage. Responsible for the
green color of pus.
NADPH-oxidase.
2O2 + NADPH ® 2O2- + NADP + H+
Key component of the respiratory burst.
Deficiency may be observed in chronic granulomatous disease.
Lysozyme.
Hydrolyzes link between N-acetylmuramic
acid and N-acetyl-D-glucosamine found in certain bacterial cell walls. Abundant
in macrophages.
Defensins.
Basic antibiotic peptides of 29-33 amino
acids. Apparently kill bacteria by causing membrane damage.
Lactoferrin.
Iron-binding protein. May inhibit growth
of certain bacteria by binding iron and may be involved in regulation of
proliferation of myeloid cells.
Neutrophils contain a number of
proteinases (elastase, collagenase, gelatinase, cathepsin G, plasminogen
activator) that can hydrolyze elastin, various types of collagens, and other
proteins present in the extracellular matrix. Such enzymatic action, if allowed
to proceed unopposed, can result in serious damage to tissues. Most of these
proteinases are lysosomal enzymes and exist mainly as inactive precursors in
normal neutrophils. Small amounts of these enzymes are released into normal
tissues, with the amounts increasing markedly during inflammation. The
activities of elastase and other proteinases are normally kept in check by a
number of antiproteinases (a1-Antiproteinase, a2-Macroglobulin, Secretory
leukoproteinase inhibitor, a1-Antichymotrypsin, Plasminogen activator
inhibitor-1, Tissue inhibitor of metalloproteinase) present in plasma and the
extracellular fluid.
Basophiles
Basophiles make up 1-5% of all blood
leukocytes. They are actively formed in the bone marrow during allergy. Basophiles take part
in the allergic reactions, in the blood
coagulation and intravascular lipolysis. They have the protein synthesis
mechanism, which works due to the biological oxidation energy . They synthesize
the mediators of allergic reactions – histamine and serotonin, which during
allergy cause local inflammation. Heparin, which is formed in the basophiles,
prevents the blood coagulation and activates intravascular lipoprotein lipase,
which splits triacylglycerin.
The number of basophils also increases
during infection. Basophils leave the blood and accumulate at the site of
infection or other inflammation. There they discharge the contents of their
granules, releasing a variety of mediators such as:
• histamine
• serotonin
• prostaglandins and
leukotrienes
which increase the blood flow to the area
and in other ways add to the inflammatory process. The mediators released by
basophils also play an important part in some allergic responses such as
• hay fever and
• an anaphylactic response to
insect stings.
Eosinophiles
They make up 3-6% of all leukocytes.
Eosinophiles as well as neutrophiles defend the cells from microorganisms, they
contain myeloperoxidase, lysosomal hydrolases. About the relations of
eosinophiles with testifies the growth of their amount during the sensitization
of organism, i.e. during bronchial asthma, helminthiasis. They are able to pile
and splits histamine, “to dissolve” thrombus with the participation of
plasminogen and bradykinin-kininase.
Monocytes
They are formed in the bone marrow. They make up 4-8% of all leukocytes.
According to the function they are called macrophages. Tissue macrophages
derive from blood monocytes. Depending on their position they are called: in
the liver – reticuloendotheliocytes, in the lungs - alveolar macrophages, in
the intermediate substance of connective tissue – histocytes etc. Monocytes are
characterized by a wide set of lysosomal
enzymes with the optimum activity in the acidic condition. The major
functions of monocytes and macrophages are endocytosis and phagocytosis.

The
amount – 20-25%, are formed in the lymphoid tissue or thymus, play important
role in the formation of humoral and cellular immunity. Lymphocytes have
powerful system of synthesis of antibody proteins, energy is majorily pertained
due to glycolysis, rarely – by aerobic way.
http://www.youtube.com/watch?v=cD_uAGPBfQQ&feature=related
There
are several kinds of lymphocytes (although they all look alike under the
microscope), each with different functions to perform . The most common types
of lymphocytes are
• B lymphocytes ("B
cells"). These are responsible for making antibodies.
• T lymphocytes ("T
cells"). There are several subsets of these:
o inflammatory T cells that
recruit macrophages and neutrophils to the site of infection or other tissue
damage
o cytotoxic T lymphocytes
(CTLs) that kill virus-infected and, perhaps, tumor cells
o helper T cells that enhance
the production of antibodies by B cells
Although
bone marrow is the ultimate source of lymphocytes, the lymphocytes that will
become T cells migrate from the bone marrow to the thymus where they mature. Both B cells and T cells
also take up residence in lymph nodes, the spleen and other tissues where they
• encounter antigens;
• continue to divide by mitosis;
• mature into fully
functional cells.
Monocytes
Monocytes
leave the blood and become macrophages and dendritic cells.
This
scanning electron micrograph (courtesy of Drs. Jan M. Orenstein and Emma
Shelton) shows a single macrophage surrounded by several lymphocytes.
Macrophages
are large, phagocytic cells that engulf
• foreign material (antigens)
that enter the body
• dead and dying cells of the
body.
Thrombocytes
(blood platelets)
Platelets
are cell fragments produced from megakaryocytes.
Blood
normally contains 150,000–350,000 per microliter (µl) or cubic millimeter
(mm3). This number is normally maintained by a homeostatic (negative-feedback)
mechanism .
The
amount – less than 1%, they play the main role in the process of hemostasis.
They are formed as a result of disintegration of megakaryocytes in the
bone marrow. Their –life-time is 7-9
days. In spite of the fact that thrombocytes have no nucleus, they are able to
perform practically all functions of the cell, besides DNA synthesis.
If
this value should drop much below 50,000/µl, there is a danger of uncontrolled
bleeding because of the essential role that platelets have in blood clotting.
Some
causes:
• certain drugs and herbal
remedies;
• autoimmunity.
When
blood vessels are cut or damaged, the loss of blood from the system must be
stopped before shock and possible death occur. This is accomplished by
solidification of the blood, a process called coagulation or clotting.
A
blood clot consists of
• a plug of platelets
enmeshed in a
• network of insoluble fibrin
molecules.
Red
Blood Cells (erythrocytes)
The
most numerous type in the blood.
• Women average about 4.8
million of these cells per cubic millimeter (mm3; which is the same as a
microliter [µl]) of blood.
• Men average about 5.4 x 106
per µl.
• These values can vary over
quite a range depending on such factors as health and altitude. (Peruvians
living at
RBC
precursors mature in the bone marrow closely attached to a macrophage.
• They manufacture hemoglobin
until it accounts for some 90% of the dry weight of the cell.
• The nucleus is squeezed out
of the cell and is ingested by the macrophage.
• No-longer-needed proteins
are expelled from the cell in vesicles called exosomes.
Human
blood contains 25 trillion of erythrocytes. Their main function –
transportation of O2 and CO2 – they perform due to the fact that they contain
34% of hemoglobin, and per dry cells mass – 95%. The total amount of hemoglobin in the blood equals
130-160 g/l. In the process of erythropoesis the preceding cells decrease their
size. Their nuclei at the end of the process are ruined and pushed out of the
cells. 90% of glucose in the erythrocytes is decomposed in the process of
glycolysis and 10% - by pentose-phosphate way. There are noted congenital
defects of enzymes of these metabolic ways of erythrocytes. During this are
usually observed hemolytic anemia and other structural and functional
erythrocytes’ affections.

This scanning
electron micrograph (courtesy of Dr. Marion J. Barnhart) shows the
characteristic biconcave shape of red blood cells.
Thus RBCs are
terminally differentiated; that is, they can never divide. They live about 120
days and then are ingested by phagocytic cells in the liver and spleen. Most of
the iron in their hemoglobin is reclaimed for reuse. The remainder of the heme
portion of the molecule is degraded intobile pigments and excreted by the
liver. Some 3 million RBCs die and are scavenged by the liver each second.
Red blood
cells are responsible for the transport of oxygen and carbon dioxide.
Oxygen
Transport
In adult
humans the hemoglobin (Hb) molecule
• consists of four
polypeptides:
o two alpha (α) chains of
141 amino acids and
o two beta (β) chains of
146 amino acids
• Each of these
is attached the prosthetic group heme.
• There is one atom of iron
at the center of each heme.
• One molecule of oxygen can
bind to each heme.
The reaction
is reversible.
• Under the conditions of
lower temperature, higher pH, and increased oxygen pressure in the capillaries
of the lungs, the reaction proceeds to the right. The purple-red deoxygenated
hemoglobin of the venous blood becomes the bright-red oxyhemoglobin of the
arterial blood.
• Under the conditions of
higher temperature, lower pH, and lower oxygen pressure in the tissues, the
reverse reaction is promoted and oxyhemoglobin gives up its oxygen.
Carbon
Dioxide Transport
Carbon
dioxide (CO2) combines with water forming carbonic acid, which dissociates into
a hydrogen ion (H+) and a bicarbonate ions

95% of the CO2 generated in the tissues is
carried in the red blood cells:
• It probably enters (and
leaves) the cell by diffusing through transmembrane channels in the plasma
membrane. (One of the proteins that forms the channel is theD antigen that is
the most important factor in the Rh system of blood groups.)
• Once inside, about one-half
of the CO2 is directly bound to hemoglobin (at a site different from the one
that binds oxygen).
• The rest is converted —
following the equation above — by the enzyme carbonic anhydrase into
o bicarbonate ions that
diffuse back out into the plasma and
o hydrogen ions (H+) that bind
to the protein portion of the hemoglobin (thus having no effect on pH).
Only about 5% of the CO2 generated in the tissues
dissolves directly in the plasma. (A good thing, too: if all the CO2 we make
were carried this way, the pH of the blood would drop from its normal 7.4 to an
instantly-fatal 4.5!)
When the red cells reach the lungs, these
reactions are reversed and CO2 is released to the air of the alveoli.

Anemia is a shortage of
• RBCs and/or
• the amount of hemoglobin in
them.
Blood
Groups
Red blood cells have surface antigens that differ
between people and that create the so-called blood groups such as the ABO
system and the Rh system.
An Essay on Hemoglobin Structure and Function:

Figure
1 is a model of human deoxyhemoglobin.
It was created in RasMol version 2.6 by Roger Sayle using the pdb
coordinates from the pdb file 4hhb. The
3D coordinates were determed from x-ray crystallography by Fermi, G., Perutz,
M. F., Shaanan, B., Fourme, R.: The crystal structure of human deoxyhaemoglobin
at
Hemoglobin is the protein that carries oxygen
from the lungs to the tissues and carries carbon dioxide from the tissues back
to the lungs. In order to function most efficiently, hemoglobin needs to bind
to oxygen tightly in the oxygen-rich atmosphere of the lungs and be able to
release oxygen rapidly in the relatively oxygen-poor environment of the
tissues. It does this in a most elegant and intricately coordinated way. The
story of hemoglobin is the prototype example of the relationship between
structure and function of a protein molecule.
Hemoglobin Structure
![]()

A
hemoglobin molecule consists of four polypeptide chains: two alpha chains, each
with 141 amino acids and two beta chains, each with 146 amino acids. The
protein portion of each of these chains is called "globin". The a and
b globin chains are very similar in structure. In this case, a and b refer to
the two types of globin. Students often confuse this with the concept of a
helix and b sheet secondary structures. But, in fact, both the a and b globin
chains contain primarily a helix secondary structure with no b sheets.

Figure 2 is a close up view of one of the heme
groups of the human a chain from dexoyhemoglobin. In this view, the iron is coordinated by a
histidine side chain from amino acid 87 (shown in green.)
Each a or b globin chain folds into
http://www.youtube.com/watch?v=eor6EK_JP40
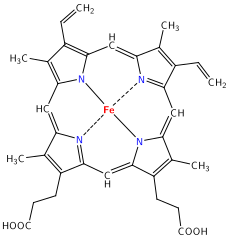
A heme group is a flat ring molecule containing
carbon, nitrogen and hydrogen atoms, with a single Fe2+ ion at the center.
Without the iron, the ring is called a porphyrin. In a heme molecule, the iron
is held within the flat plane by four nitrogen ligands from the porphyrin ring.
The iron ion makes a fifth bond to a histidine side chain from one of the
helices that form the heme pocket. This fifth coordination bond is to histidine
The Bohr Effect
The ability of hemoglobin to release oxygen, is
affected by pH, CO2 and by the differences in the oxygen-rich environment of
the lungs and the oxygen-poor environment of the tissues. The pH in the tissues
is considerably lower (more acidic) than in the lungs. Protons are generated
from the reaction between carbon dioxide and water to form bicarbonate:
CO2 + H20 -----------------> HCO3- + H+
This increased acidity serves a twofold purpose.
First, protons lower the affinity of hemoglobin for oxygen, allowing easier
release into the tissues. As all four oxygens are released, hemoglobin binds to
two protons. This helps to maintain equilibrium towards the right side of the
equation. This is known as the Bohr effect, and is vital in the removal of
carbon dioxide as waste because CO2 is insoluble in the bloodstream. The
bicarbonate ion is much more soluble, and can thereby be transported back to
the lungs after being bound to hemoglobin. If hemoglobin couldn’t absorb the
excess protons, the equilibrium would shift to the left, and carbon dioxide
couldn’t be removed.
In the lungs, this effect works in the reverse
direction. In the presence of the high oxygen concentration in the lungs, the
proton affinity decreases. As protons are shed, the reaction is driven to the
left, and CO2 forms as an insoluble gas to be expelled from the lungs. The
proton poor hemoglobin now has a greater affinity for oxygen, and the cycle
continues.
Haemoglobin or hemoglobin (frequently abbreviated
as Hb or Hgb) is the iron-containing oxygen-transport metalloprotein in the red
blood cells of the blood in vertebratesand other animals; in mammals the
protein makes up about 97% of the red cell’s dry content, and around 35% of the
total content including water. Hemoglobin transports oxygen from the lungs or
gills to the rest of the body, such as to the muscles, where it releases the
oxygen load. Hemoglobin also has a variety of other gas-transport and
effect-modulation duties, which vary from species to species, and which in
invertebrates may be quite diverse.
The name hemoglobin is the concatenation of heme
and globin, reflecting the fact that each subunit of hemoglobin is a globular
protein with an embedded heme (or haem) group; each heme group contains an iron
atom, and this is responsible for the binding of oxygen. The most common type
of hemoglobin in mammals contains four such subunits, each with one heme group.
Mutations in the genes for the hemoglobin protein
in humans result in a group of hereditary diseases termed the hemoglobinopathies,
the most common members of which are sickle-cell disease and thalassemia.
Historically in human medicine, the hemoglobinopathy of sickle-cell disease was
the first disease to be understood in its mechanism of dysfunction, completely
down to the molecular level. However, not all of such mutations produce disease
states, and are formally recognized as hemoglobin variants (not diseases).

Hemoglobin (Hb) is synthesized in a complex
series of steps. The heme portion is sythesized in both the the mitochondria
and cytosol of the immature red blood cell, while the globin protein portions
of the molecule are sythesized by ribosomes in the cytosol [3]. Production of
Hb continues in the cell throughout its early development from
theproerythroblast to the reticulocyte in the bone marrow. At this point, the
nucleus is lost in mammals, but not in birds and many other species. Even after
the loss of the nucleus in mammals, however, residual ribosomal RNA allows
further synthesis of Hb until the reticulocyte loses its RNA soon after
entering the vasculature (this hemoglobin-synthetic RNA in fact gives the
reticulocyte its reticulated appearance and name).
The empirical chemical formula of the most common
human hemoglobin is C2952H4664N812O832S8Fe4, but as noted above, hemoglobins
vary widely across species, and even (through common mutations) slightly among
subgroups of humans.
In humans, the hemoglobin molecule is an assembly
of four globular protein subunits. Each subunit is composed of a protein chain
tightly associated with a non-proteinheme group. Each protein chain arranges
into a set of alpha-helix structural segments connected together in a globin
fold arrangement, so called because this arrangement is the same folding motif
used in other heme/globin proteins such as myoglobin. This folding pattern
contains a pocket which strongly binds the heme group.
A heme group consists of an iron (Fe) atom held
in a heterocyclic ring, known as a porphyrin. The iron atom, which is the site
of oxygen binding, bonds with the fournitrogens in the center of the ring,
which all lie in one plane. The iron is also bound strongly to the globular
protein via the imidazole ring of a histidine residue below the porphyrin ring.
A sixth position can reversibly bind oxygen, completing the octahedral group of
six ligands. Oxygen binds in an "end-on bent" geometry where one
oxygen atom binds Fe and the other protrudes at an angle. When oxygen is not
bound, a very weakly bonded water molecule fills the site, forming a distorted
octahedron.
The iron atom may either be in the Fe2+ or Fe3+
state, but ferrihemoglobin (methemoglobin) (Fe3+) cannot bind oxygen. In
binding, oxygen temporarily oxidizes Fe to (Fe3+), so iron must exist in the +2
oxidation state in order to bind oxygen. The body reactivates hemoglobin found
in the inactive (Fe3+) state by reducing the iron center.
In adult humans, the most common hemoglobin type
is a tetramer (which contains 4 subunit proteins) called hemoglobin A,
consisting of two α and two β subunits non-covalently bound, each
made of 141 and 146 amino acid residues, respectively. This is denoted as
α2β2. The subunits are structurally similar and about the same size.
Each subunit has a molecular weight of about 17,000 daltons, for a total
molecular weight of the tetramer of about 68,000 daltons. Hemoglobin A is the
most intensively studied of the hemoglobin molecules.
The four polypeptide chains are bound to each
other by salt bridges, hydrogen bonds, and hydrophobic interactions. There are
two kinds of contacts between the α and β chains: α1β1 and
α1β2.
Oxyhemoglobin is formed during respiration when
oxygen binds to the heme component of the protein hemoglobin in red blood
cells. This process occurs in the pulmonary capillaries adjacent to the alveoli
of the lungs. The oxygen then travels through the blood stream to be dropped
off at cells where it is utilized in aerobic glycolysis and in the production
of ATP by the process of oxidative phosphorylation. It doesn't however help to
counteract a decrease in blood pH. Ventilation, or breathing, may reverse this
condition by removal of carbon dioxide, thus causing a shift up in pH.
Deoxyhemoglobin is the form of hemoglobin without
the bound oxygen. The absorption spectra of oxyhemoglobin and deoxyhemoglobin
differ. The oxyhemoglobine has significantly lower absorption of the 660 nm
wavelength than deoxyhemoglobin, while at 940 nm its absorption is slightly
higher. This difference is used for measurement of the amount of oxygen in
patient's blood by an instrument called pulse oximeter.


Iron's oxidation state in oxyhemoglobin
The oxidation state of iron in hemoglobin is
always +2. It does not change when oxygen binds to the deoxy- form.
Assigning oxygenated hemoglobin's oxidation state
is difficult because oxyhemoglobin is diamagnetic (no net unpaired electrons),
but the low-energy electron configurations in both oxygen and iron are
paramagnetic. Triplet oxygen, the lowest energy oxygen species, has two
unpaired electrons in antibonding π* molecular
orbitals. Iron(II) tends to be in a high-spin configuration where unpaired
electrons exist in eg antibonding orbitals. Iron(III) has an odd number of
electrons and necessarily has unpaired electrons. All of these molecules are
paramagnetic (have unpaired electrons), not diamagnetic, so an unintuitive
distribution of electrons must exist to induce diamagnetism.
The three logical possibilities are:
1) Low-spin Fe2+ binds to high-energy singlet
oxygen. Both low-spin iron and singlet oxygen are diamagnetic.
2) High-spin Fe3+ binds to .O2- (the superoxide
ion) and antiferromagnetism oppositely aligns the two unpaired electrons,
giving diamagnetic properties.
3) Low-spin Fe4+ binds to O22-. Both are
diamagnetic.
X-ray photoelectron spectroscopy suggests that
iron has an oxidation state of approximately 3.2 and infrared stretching
frequencies of the O-O bond suggests a bond length fitting with superoxide. The
correct oxidation state of iron is thus the +3 state with oxygen in the -1
state. The diamagnetism in this configuration arises from the unpaired electron
on superoxide aligning antiferromagnetically in the opposite direction from the
unpaired electron on iron. The second choice being correct is not surprising
because singlet oxygen and large separations of charge are both unfavorably
high-energy states. Iron's shift to a higher oxidation state decreases the
atom's size and allows it into the plane of the porphyrin ring, pulling on the
coordinated histidine residue and initiating the allosteric changes seen in the
globulins. The assignment of oxidation state, however, is only a formalism so
all three models may contribute to some small degree.
Early postulates by bioinorganic chemists claimed
that possibility (1) (above) was correct and that iron should exist in
oxidation state II (indeed iron oxidation state III as methemoglobin, when not
accompanied by superoxide .O2- to "hold" the oxidation electron, is
incapable of binding O2). The iron chemistry in this model was elegant, but the
presence of singlet oxygen was never explained. It was argued that the binding
of an oxygen molecule placed high-spin iron(II) in an octahedral field of
strong-field ligands; this change in field would increase the crystal field
splitting energy, causing iron's electrons to pair into the diamagnetic
low-spin configuration.
Binding of ligands
Binding and release of ligands induces a
conformational (structural) change in hemoglobin. Here, the binding and release
of oxygen illustrates the structural differences between oxy- and
deoxyhemoglobin, respectively. Only one of the four heme groups is shown.
As discussed above, when oxygen binds to the iron
center it causes contraction of the iron atom, and causes it to move back into
the center of the porphyrin ring plane (see moving diagram). At the same time,
the porphyrin ring plane itself is pushed away from the oxygen and toward the imidizole
side chain of the histidine residue interacting at the other pole of the iron.
The interaction here forces the ring plane sideways toward the outside of the
tetramer, and also induces a strain on the protein helix containing the
histidine, as it moves nearer the iron. This causes a tug on this peptide
strand which tends to open up heme units in the remainder of the molecule, so
that there is more room for oxygen to bind at their heme sites.
In the tetrameric form of normal adult
hemoglobin, the binding of oxygen is thus a cooperative process. The binding
affinity of hemoglobin for oxygen is increased by the oxygen saturation of the
molecule, with the first oxygens bound influencing the shape of the binding
sites for the next oxygens, in a way favorable for binding. This positive
cooperative binding is achieved through steric conformational changes of the
hemoglobin protein complex as discussed above, i.e. when one subunit protein in
hemoglobin becomes oxygenated, this induces a conformational or structural
change in the whole complex, causing the other subunits to gain an increased
affinity for oxygen. As a consequence, the oxygen binding curve of hemoglobin
is sigmoidal, or S-shaped, as opposed to the normal hyperbolic curve associated
with noncooperative binding.
Hemoglobin's oxygen-binding capacity is decreased
in the presence of carbon monoxide because both gases compete for the same
binding sites on hemoglobin, carbon monoxide binding preferentially in place of
oxygen. Carbon dioxide occupies a different binding site on the hemoglobin.
Through the enzyme carbonic anhydrase, carbon dioxide reacts with water to give
carbonic acid, which decomposes into bicarbonate and protons:
CO2 + H2O → H2CO3 → HCO3- + H+
The
sigmoidal shape of hemoglobin's oxygen-dissociation curve results from
cooperative binding of oxygen to hemoglobin.
Hence
blood with high carbon dioxide levels is also lower in pH (more acidic).
Hemoglobin can bind protons and carbon dioxide which causes a conformational
change in the protein and facilitates the release of oxygen. Protons bind at
various places along the protein, and carbon dioxide binds at the alpha-amino
group forming carbamate. Conversely, when the carbon dioxide levels in the
blood decrease (i.e., in the lung capillaries), carbon dioxide and protons are
released from hemoglobin, increasing the oxygen affinity of the protein. This
control of hemoglobin's affinity for oxygen by the binding and release of
carbon dioxide and acid, is known as the Bohr effect.
![]()

The binding of oxygen is affected by molecules such as carbon monoxide
(CO) (for example from tobacco smoking, cars and furnaces). CO competes with
oxygen at the heme binding site. Hemoglobin binding affinity for CO is 200
times greater than its affinity for oxygen, meaning that small amounts of CO
dramatically reduces hemoglobin's ability to transport oxygen. When hemoglobin
combines with CO, it forms a very bright red compound called carboxyhemoglobin.
When inspired air contains CO levels as low as 0.02%, headache and nausea
occur; if the CO concentration is increased to 0.1%, unconsciousness will
follow. In heavy smokers, up to 20% of the oxygen-active sites can be blocked
by CO.
In similar fashion, hemoglobin also has competitive binding affinity for
cyanide (CN-), sulfur monoxide (SO), nitrogen dioxide (NO2), and sulfide (S2-),
including hydrogen sulfide (H2S). All of these bind to iron in heme without
changing its oxidation state, but they nevertheless inhibit oxygen-binding,
causing grave toxicity.
The iron atom in the heme group must be in the Fe2+ oxidation state to
support oxygen and other gases' binding and transport. Oxidation to Fe3+ state
converts hemoglobin into hemiglobin or methemoglobin (pronounced
"MET-hemoglobin"), which cannot bind oxygen. Hemoglobin in normal red
blood cells is protected by a reduction system to keep this from happening.
Nitrogen dioxide and nitrous oxide are capable of converting a small fraction
of hemoglobin to methemoglobin, however this is not usually of medical
importance (nitrogen dioxide is poisonous by other mechanisms, and nitrous
oxide is routinely used in surgical anesthesia in most people without undue
methemoglobin buildup).
In people acclimated to high altitudes, the concentration of
2,3-bisphosphoglycerate (2,3-BPG) in the blood is increased, which allows these
individuals to deliver a larger amount of oxygen to tissues under conditions of
lower oxygen tension. This phenomenon, where molecule Y affects the binding of
molecule X to a transport molecule Z, is called a heterotropic allosteric
effect.
A variant hemoglobin, called fetal hemoglobin (HbF, α2γ2), is
found in the developing fetus, and binds oxygen with greater affinity than
adult hemoglobin. This means that the oxygen binding curve for fetal hemoglobin
is left-shifted (i.e., a higher percentage of hemoglobin has oxygen bound to it
at lower oxygen tension), in comparison to that of adult hemoglobin. As a
result, fetal blood in the placenta is able to take oxygen from maternal blood.
Hemoglobin also carries nitric oxide in the globin part of the molecule.
This improves oxygen delivery in the periphery and contributes to the control
of respiration. NO binds reversibly to a specific cystein residue in globin;
the binding depends on the state (R or T) of the hemoglobin. The resulting
S-nitrosylated hemoglobin influences various NO-related activities such as the
control of vascular resistance, blood pressure and respiration. NO is released
not in the cytoplasm of erythrocytes but is transported by an anion exchanger
called AE1 out of them.[]
Degradation of hemoglobin in vertebrate animals
When red cells reach the end of their life due to aging or defects, they
are broken down, the hemoglobin molecule is broken up and the iron gets
recycled. When the porphyrin ring is broken up, the fragments are normally
secreted in the bile by the liver. This process also produces one molecule of
carbon monoxide for every molecule of heme degraded [4]; this is one of the few
natural sources of carbon monoxide production in the human body, and is
responsible for the normal blood levels of carbon monoxide even in people
breathing pure air. The other major final product of heme degradation is
bilirubin. Increased levels of this chemical are detected in the blood if red
cells are being destroyed more rapidly than usual. Improperly degraded
hemoglobin protein or hemoglobin that has been released from the blood cells
too rapidly can clog small blood vessels, especially the delicate blood
filtering vessels of the kidneys, causing kidney damage
Role in disease
Decrease of hemoglobin, with or without an absolute decrease of red
blood cells, leads to symptoms of anemia. Anemia has many different causes,
although iron deficiencyand its resultant iron deficiency anemia are the most
common causes in the Western world. As absence of iron decreases heme
synthesis, red blood cells in iron deficiency anemia are hypochromic (lacking
the red hemoglobin pigment) and microcytic (smaller than normal). Other anemias
are rarer. In hemolysis (accelerated breakdown of red blood cells), associated
jaundice is caused by the hemoglobin metabolite bilirubin, and the circulating
hemoglobin can cause renal failure.
Some mutations in the globin chain are associated with the
hemoglobinopathies, such as sickle-cell disease and thalassemia. Other
mutations, as discussed at the beginning of the article, are benign and are
referred to merely as hemoglobin variants.
There is a group of genetic disorders, known as the porphyrias that are
characterized by errors in metabolic pathways of heme synthesis. King George
III of the
To a small extent, hemoglobin A slowly combines with glucose at a
certain location in the molecule. The resulting molecule is often referred to
as Hb A1c. As theconcentration of glucose in the blood increases, the
percentage of Hb A that turns into Hb A1c increases. In diabetics whose glucose
usually runs high, the percent Hb A1c also runs high. Because of the slow rate
of Hb A combination with glucose, the Hb A1c percentage is representative of
glucose level in the blood averaged over a longer time (the half-life of red
blood cells, which is typically 50-55 days).
Diagnostic use
Hemoglobin levels are amongst the most commonly performed blood tests,
usually as part of a full blood count or complete blood count. Results are
reported in g/L, g/dLor mol/L. For conversion, 1 g/dL is 0.621 mmol/L. If the
total hemoglobin concentration in the blood falls below a set point, this is
called anemia. Normal values for hemoglobin levels are:
• Women: 12.1 to 15.1
g/dl
• Men: 13.8 to 17.2
g/dl
• Children: 11 to 16
g/dl
• Pregnant women: 11
to 12 g/dl
Anemias are further subclassified by the size of the red blood cells,
which are the cells which contain hemoglobin in vertebrates. They can be
classified as microcytic (small sized red blood cells), normocytic (normal
sized red blood cells), or macrocytic (large sized red blood cells). The
hemaglobin is the typical test used for blood donation. A comparison with the
hematocrit can be made by multiplying the hemaglobin by three. For example, if
the hemaglobin is measured at 17, that compares with a hematocrit of .51
Glucose levels in blood can vary widely each hour, so one or only a few
samples from a patient analyzed for glucose may not be representative of
glucose control in the long run. For this reason a blood sample may be analyzed
for Hb A1c level, which is more representative of glucose control averaged over
a longer time period (determined by the half-life of the individual's red blood
cells, which is typically 50-55 days). People whose Hb A1c runs 6.0% or less
show good longer-term glucose control. Hb A1c values which are more than 7.0%
are elevated. This test is especially useful for diabetics.[8]
BIOLOGICAL BUFFERS OF BLOOD
Acid–base homeostasis is the part of human homeostasis concerning the
proper balance between acids and bases, also called body pH. The body is very
sensitive to its pH level, so strong mechanisms exist to maintain it. Outside
the acceptable range of pH, proteins are denatured and digested, enzymes lose
their ability to function, and death may occur.
A buffer solution is an aqueous solution consisting of a mixture of a
weak acid and its conjugate base or a weak base and its conjugate acid. Its pH
changes very little when a small amount of strong acid or base is added to it.
Buffer solutions are used as a means of keeping pH at a nearly constant value
in a wide variety of chemical applications.
Many life forms thrive only in a relatively small pH range so they
utilize a buffer solution to maintain a constant pH. One example of a buffer
solution found in nature is blood. The body's acid–base balance is normally
tightly regulated, keeping the arterial blood pH between 7.38 and 7.42. Several
buffering agents that reversibly bind hydrogen ions and impede any change in pH
exist. Extracellular buffers include bicarbonate and ammonia, whereas proteins
and phosphate act as intracellular buffers. The bicarbonate buffering system is
especially key, as carbon dioxide (CO2) can be shifted through carbonic acid
(H2CO3) to hydrogen ions and bicarbonate (HCO3-):
Acid–base imbalances that overcome the buffer system can be compensated
in the short term by changing the rate of ventilation. This alters the
concentration of carbon dioxide in the blood, shifting the above reaction
according to Le Chatelier's principle, which in turn alters the pH.
The kidneys are slower to compensate, but renal physiology has several
powerful mechanisms to control pH by the excretion of excess acid or base. In
response to acidosis, tubular cells reabsorb more bicarbonate from the tubular
fluid, collecting duct cells secrete more hydrogen and generate more
bicarbonate, and ammoniagenesis leads to increased formation of the NH3 buffer.
In responses to alkalosis, the kidneys may excrete more bicarbonate by
decreasing hydrogen ion secretion from the tubular epithelial cells, and
lowering rates of glutamine metabolism and ammonium excretion.
Buffering hides from view the
real change in H+ that occurs.
This huge buffer capacity has another not immediately obvious
implication for how we think about the severity of an acid-base disorder. You
would think that the magnitude of an acid-base disturbance could be quantified
merely by looking at the change in [H+] - BUT this is not so.
Because of the large buffering capacity, the actual change in [H+] is so
small it can be ignored in any quantitative assessment, and instead, the
magnitude of a disorder has to be estimated indirectly from the decrease in the
total concentration of the anions involved in the buffering. The buffer anions,
represented as A-, decrease because they combine stoichiometrically with H+ to
produce HA. A decrease in A- by 1 mmol/l represents a 1,000,000 nano-mol/l
amount of H+ that is hidden from view and this is several orders of magnitude
higher than the visible few nanomoles/l change in [H+] that is visible.) - As
noted above in the comments about the Swan & Pitts experiment, 13,999,994
out of 14,000,000 nano-moles/l of H+ were hidden on buffers and just to count
the 36 that were on view would give a false impression of the magnitude of the
disorder.
The Bicarbonate Buffer System
The major buffer system in the
ECF is the CO2-bicarbonate buffer system. This is responsible for about 80% of
extracellular buffering. It is the most important ECF buffer for metabolic
acids but it cannot buffer respiratory acid-base disorders.
The components are easily
measured and are related to each other by the Henderson-Hasselbalch equation.
Henderson-Hasselbalch Equation
pH = pK’a + log10 ( [HCO3] /
0.03 x pCO2)
The pK’a value is dependent on
the temperature, [H+] and the ionic concentration of the solution. It has a
value of 6.099 at a temperature of 37C
and a plasma pH of 7.4. At a temperature of 30C and pH of 7.0, it has a value of 6.148. For
practical purposes, a value of 6.1 is generally assumed and corrections for
temperature, pH of plasma and ionic strength are not used except in precise
experimental work.
The pK'a is derived from the
Ka value of the following reaction:
CO2 + H2O <=> H2CO3
<=> H+ + HCO3-
(where CO2 refers to dissolved
CO2)
The concentration of carbonic
acid is very low compared to the other components so the above equation is
usually simplified to:
CO2 + H2O <=> H+ + HCO3-
By the Law of Mass Action:
Ka = [H+] . [HCO3-] / [CO2] .
[H20]
The concentration of H2O is so
large (55.5M) compared to the other components, the small loss of water due to this
reaction changes its concentration by only an extremely small amount. This
means that [H2O] is effectively constant. This allows further simplification as
the two constants (Ka and [H2O] ) can be combined into a new constant K’a.
K’a = Ka x [H2O] = [H+] .
[HCO3-] / [CO2]
Substituting:
K'a = 800 nmol/l (value for
plasma at 37C )
[CO2] = 0.03 x pCO2 (by
Henry’s Law) [where 0.03 is the solubility coefficient]
into the equation yields the
Henderson Equation:
[H+] = (800 x 0.03) x pCO2 /
[HCO3-] = 24 x pCO2 / [HCO3-] nmol/l
Taking the logs (to base 10)
of both sides yields the Henderson-Hasselbalch equation:
pH = log10(800) - log (0.03
pCO2 / [HCO3-] )
pH = 6.1 + log ( [HCO3] / 0.03
pCO2 )
On chemical grounds, a
substance with a pKa of 6.1 should not be a good buffer at a pH of 7.4 if it
were a simple buffer. The system is more complex as it is ‘open at both ends’
(meaning both [HCO3] and pCO2 can be adjusted) and this greatly increases the
buffering effectiveness of this system. The excretion of CO2 via the lungs is
particularly important because of the rapidity of the response. The adjustment
of pCO2 by change in alveolar ventilation has been referred to as physiological
buffering.
The bicarbonate buffer system
is an effective buffer system despite having a low pKa because the body also
controls pCO2
Other Buffers
The other buffer systems in
the blood are the protein and phosphate buffer systems.
These are the only blood
buffer systems capable of buffering respiratory acid-base disturbances as the
bicarbonate system is ineffective in buffering changes in H+ produced by
itself.
The phosphate buffer system is
NOT an important blood buffer as its concentration is too low
The concentration of phosphate
in the blood is so low that it is quantitatively unimportant. Phosphates are
important buffers intracellularly and in urine where their concentration is
higher.
Phosphoric acid is triprotic
weak acid and has a pKa value for each of the three dissociations:
The three pKa values are sufficiently different so that at any one pH
only the members of a single conjugate pair are present in significant
concentrations.
At the prevailing pH values in most biological systems, monohydrogen
phosphate (HPO4-2) and dihydrogen phosphate (H2PO4-) are the two species
present. The pKa2 is 6.8 and this makes the closed phosphate buffer system a
good buffer intracellularly and in urine. The pH of glomerular ultrafiltrate is
7.4 and this means that phosphate will initially be predominantly in the
monohydrogen form and so can combine with more H+ in the renal tubules. This
makes the phosphate buffer more effective in buffering against a drop in pH
than a rise in pH.
Note: The ‘true’ pKa2 value is actually 7.2 if measured at zero ionic
strength but at the typical ionic strength found in the body its apparent value
is 6.8. The other factor which makes phosphate a more effective buffer
intracellularly and in urine is that its concentration is much higher here than
in extracellular fluid.
Haemoglobin is an important blood buffer particularly for buffering CO2
Protein buffers in blood include haemoglobin (150g/l) and plasma
proteins (70g/l). Buffering is by the imidazole group of the histidine residues
which has a pKa of about 6.8. This is suitable for effective buffering at
physiological pH. Haemoglobin is quantitatively about 6 times more important then
the plasma proteins as it is present in about twice the concentration and
contains about three times the number of histidine residues per molecule. For
example if blood pH changed from 7.5 to 6.5, haemoglobin would buffer 27.5
mmol/l of H+ and total plasma protein buffering would account for only 4.2
mmol/l of H+.
Deoxyhaemoglobin is a more effective buffer than oxyhaemoglobin and this
change in buffer capacity contributes about 30% of the Haldane effect. The
major factor accounting for the Haldane effect in CO2 transport is the much
greater ability of deoxyhaemoglobin to form carbamino compounds.
This buffer functions in exactly the same way as the phosphate buffer.
Additional H+ is consumed by HCO3- and additional
The concentrations of
hydrogen carbonate ions and of carbonic acid are controlled by two independent
physiological systems. Carbonic acid concentration is controlled by
respiration, that is through the lungs. Carbonic acid is in equilibrium with
dissolved carbon dioxide gas.
H2CO3(aq) CO2(aq) + H2O(l)
An enzyme called carbonic anhydrase catalyzes the conversion of carbonic
acid to dissolved carbon dioxide. In the lungs, excess dissolved carbon dioxide
is exhaled as carbon dioxide gas.
CO2(aq) CO2(g)
The concentration of hydrogen carbonate ions is controlled through the
kidneys. Excess hydrogen carbonate ions are excreted in the urine.
The much higher concentration
of hydrogen carbonate ion over that of carbonic acid in blood plasma allows the
buffer to respond effectively to the most common materials that are released
into the blood. Normal metabolism releases mainly acidic materials: carboxylic
acids such as lactic acid (HLac). These acids react with hydrogen carbonate ion
and form carbonic acid.
HLac(aq) + HCO3-(aq) Lac-(aq) +
H2CO3(aq)
The carbonic acid is converted through the action of the enzyme carbonic
anhydrase into aqueous carbon dioxide.
H2CO3(aq) CO2(aq) + H2O(l)
An increase in CO2(aq) concentration stimulates increased breathing, and
the excess carbon dioxide is released into the air in the lungs.
The condition called
respiratory acidosis occurs when blood pH falls as a result of decreased
respiration. When respiration is restricted, the concentration of dissolved
carbon dioxide in the blood increases, making the blood too acidic. Such a
condition can be produced by asthma, pneumonia, emphysema, or inhaling smoke.
Metabolic acidosis is the
decrease in blood pH that results when excessive amounts of acidic substances
are released into the blood. This can happen through prolonged physical
exertion, by diabetes, or restricted food intake. The normal body response to
this condition is increases breathing to reduce the amount of dissolved carbon
dioxide in the blood. This is why we breathe more heavily after climbing several
flights of stairs.
Respiratory alkalosis results
from excessive breathing that produces an increase in blood pH.
Hyperventilation causes too much dissolved carbon dioxide to be removed from
the blood, which decreases the carbonic acid concentration, which raises the
blood pH. Often, the body of a hyperventilating person will react by fainting,
which slows the breathing.
Metabolic alkalosis is an
increase in blood pH resulting from the release of alkaline materials into the
blood. This can result from the ingestion of alkaline materials, and through
overuse of diuretics. Again, the body usually responds to this condition by
slowing breathing, possibly through fainting.
The carbonic acid-hydrogen
carbonate ion buffer works throughout the body to maintain the pH of blood
plasma close to 7.40. The body maintains the buffer by eliminating either the
acid (carbonic acid) or the base (hydrogen carbonate ions). Changes in carbonic
acid concentration can be effected within seconds through increased or
decreased respiration. Changes in hydrogen carbonate ion concentration,
however, require hours through the relatively slow elimination through the
kidneys

Plasma
Plasma is the straw-colored liquid in which the blood cells
are suspended.
Plasma transports materials needed by cells and
materials that must be removed from cells:
• various ions (Na+, Ca2+,
HCO3−, etc.
• glucose and traces of other
sugars
• amino acids
• other organic acids
• cholesterol and other
lipids
• hormones
• urea and other wastes
Most of these materials are in transit from a
place where they are added to the blood (a "source")
• exchange organs like the
intestine
• depots of materials like
the liver
to places ("sinks") where they will be
removed from the blood.
• every cell
• exchange organs like the
kidney, and skin.
Serum Proteins
Proteins make up 6–8% of the blood. They are
about equally divided between serum albumin and a great variety of serum
globulins.
After blood is withdrawn from a vein and allowed
to clot, the clot slowly shrinks. As it does so, a clear fluid called serum is
squeezed out. Thus:
Serum is blood plasma without fibrinogen and
other clotting factors.
The serum proteins can be separated by
electrophoresis.
• A drop
of serum is applied in a band to a thin sheet of supporting material, like
paper, that has been soaked in a slightly-alkaline salt solution.
• At pH
8.6, which is commonly used, all the proteins are negatively charged, but some
more strongly than others.
• A
direct current can flow through the paper because of the conductivity of the
buffer with which it is moistened.
• As the
current flows, the serum proteins move toward the positive electrode.
• The
stronger the negative charge on a protein, the faster it migrates.
• After a
time (typically 20 min), the current is turned off and the proteins stained to
make them visible (most are otherwise colorless).
• The
separated proteins appear as distinct bands.
• The
most prominent of these and the one that moves closest to the positive
electrode is serum albumin.
• Serum
albumin
o is
made in the liver
o
binds many small molecules for transport through the blood
o
helps maintain the osmotic pressure of the blood
• The
other proteins are the various serum globulins.
• They
migrate in the order
o
alpha globulins (e.g., the proteins that transport thyroxine and retinol
[vitamin A])
o beta
globulins (e.g., the iron-transporting protein transferrin)
o
gamma globulins.
§
Gamma globulins are the least negatively-charged serum proteins. (They
are so weakly charged, in fact, that some are swept in the flow of buffer back
toward the negative electrode.)
§
Most antibodies are gamma globulins.
§
Therefore gamma globulins become more abundant following infections or
immunizations.
Albumins – multidispersed fraction of blood
plasma which are characterized by the high electrophoretic mobility and mild
dissolubility in water and saline solutions. Molecular weight of albumins is
about 60000. Due to high hydrophilic properties albumins bind a significant
amount of water, and the volume of their molecule under hydratation is doubled.
Hydrative layer formed around the serum albumins provides to 70-80 % of oncotic
pressure of blood plasma proteins, that can be applied in clinical practice at
albumins transfusion to patients with tissue edemas. The decreasing of albumins
concentration in blood plasma, for example under disturbance of their synthesis
in hepatocytes at liverfailure, can cause the water transition from a vessels
into the tissues and development of oncotic edemas.
Albumins execute also important physiological
function as transporters of a lot of metabolites and diverse low molecular
weight structures. The molecules of albumins have several sites with centers of
linkage for molecules of organic ligands, which are affixed by the
electrostatic and hydrophobic bonds. Serum albumins can affix and convey fatty
acids, cholesterol, cholic pigments (bilirubin and that similar), vitamins,
hormones, some amino acids, toxins and medicines.
Albumins also execute the buffer function. Due to
the availability in their structure amino and carboxylic groups albumins can
react both as acids and as alkaline.
Albumins can bound different toxins in blood
plasma (bilirubin, foreign substances et c.). This is the desintoxicative function of albumins.
Albumins also play role of amino acids depot in
the organism. They can supply amino acids for the building of another proteins,
for example enzymes.
Globulins - heterogeneous fraction of blood
proteins which execute transport (a1-globulins – transport of lipids, thyroxin,
corticosteroid hormones; a2-globulins - transport of lipids, copper ions;
b-globulins - transport of lipids, iron) and protective (participation of
b-globulins in immune reactions as antitoxins; g-globulins as immunoglobulins)
functions. They also support the blood oncotic pressure and acid-alkaline
balance, provide amino acids for the organism requirements. The molecular
weight of globulins is approximately 150000-300000.
The globulin level in blood plasma is 20-40 g/l.
A ratio between concentrations of albumins and globulins (so called “protein
coefficient”) in blood plasma is often determined in clinical practice. In
healthy people this coefficient is 1,5-2,0.
Fibrinogen – important protein of blood plasma,
precursor of fibrin, the structural element of blood clots. Fibrinogen
participates in blood clotting and thus prevents the loss of blood from the
vascular system of vertebrates. The approximate molecular weight of fibrinogen
is 340000. It is the complex protein, it contains the carbohydrate as
prosthetic group. The content of firinogen in blood is 3-4 g/l.
Subfractions of a1, a2, b and g globulins, their
structure and functions.
Immunoglobulins
(Ig A, Ig G, Ig E, Ig M) - proteins of g-globulin fraction of blood plasma
executing the functions of antibodies which are the main effectors of humoral
immunity. They appear in the blood serum and certain cells of a vertebrate in
response to the introduction of a protein or some other macromolecule foreign
to that species.
Immunoglobulin molecules have bindind sites that
are specific for and complementary to the structural features of the antigen
that induced their formation. Antibodies are highly specific for the foreign
proteins that evoke their formation.
Molecules of immunoglobulins are glycoproteins. The protein part of
immunoglobulins contain four polipeptide
chains: two heavy H-chains and two light L-chains.
The acute phase response develops in a wide range
of acute and chronic inflammatory conditions like bacterial, viral, or fungal
infections; rheumatic and other inflammatory diseases; malignancy; and tissue
injury or necrosis. These conditions cause release of interleukin-6 and other
cytokines that trigger the synthesis of CRP and fibrinogen by the liver. During
the acute phase response, levels of CRP rapidly increase within 2 hours of
acute insult, reaching a peak at 48 hours. With resolution of the acute phase
response, CRP declines with a relatively short half-life of 18 hours. Measuring
CRP level is a screen for infectious and inflammatory diseases. Rapid, marked
increases in CRP occur with inflammation, infection, trauma and tissue
necrosis, malignancies, and autoimmune disorders. Because there are a large
number of disparate conditions that can increase CRP production, an elevated
CRP level does not diagnose a specific disease. An elevated CRP level can
provide support for the presence of an inflammatory disease, such as rheumatoid
arthritis, polymyalgia rheumatica or giant-cell arteritis.
The physiological role of CRP is to bind to
phosphocholine expressed on the surface of dead or dying cells (and some types
of bacteria) in order to activate the complement system. CRP binds to
phosphocholine on microbes and damaged cells and enhances phagocytosis by
macrophages. Thus, CRP participates in the clearance of necrotic and apoptotic
cells.
CRP is a member of the class of acute-phase
reactants, as its levels rise dramatically during inflammatory processes
occurring in the body. This increment is due to a rise in the plasma
concentration of IL-6, which is produced predominantly by macrophages[2] as
well asadipocytes. CRP binds to phosphocholine on microbes. It is thought to
assist in complement binding to foreign and damaged cells and enhances
phagocytosis by macrophages (opsonin mediated phagocytosis), which express a
receptor for CRP. It is also believed to play another important role in innate
immunity, as an early defense system against infections.
CRP rises up to 50,000-fold in acute
inflammation, such as infection. It rises above normal limits within 6 hours,
and peaks at 48 hours. Its half-life is constant, and therefore its level is
mainly determined by the rate of production (and hence the severity of the
precipitating cause).
Serum amyloid A is a related acute-phase marker
that responds rapidly in similar circumstances.

In blood plasma of healthy people the C-reactive
protein is absent but it occurs at pathological states accompanied by an inflammation
and necrosis of tissues. The availability of C-reactive protein is
characteristic for the acute period of diseases – “protein of an acute phase”.
The determination of C-reactive protein has diagnostic value in an acute phase
of rheumatic disease, at a myocardial infarction, pneumococcal, streptococcal,
staphylococcal infections.
Diagnostic use
CRP is used mainly as a marker of inflammation.
Apart from liver failure, there are few known factors that interfere with CRP
production.[2]
Measuring and charting CRP values can prove
useful in determining disease progress or the effectiveness of treatments.
Blood, usually collected in a serum-separating tube, is analysed in amedical
laboratory or at the point of care. Various analytical methods are available
for CRP determination, such as ELISA, immunoturbidimetry, rapid
immunodiffusion, and visual agglutination.
Reference ranges for blood tests, showing
C-reactive protein in brown-yellow in center.
A high-sensitivity CRP (hs-CRP) test measures low
levels of CRP using laser nephelometry. The test gives results in 25 minutes
with a sensitivity down to 0.04 mg/L.
Normal concentration in healthy human serum is
usually lower than 10 mg/L, slightly increasing with aging. Higher levels are
found in late pregnant women, mild inflammation and viral infections (10–40
mg/L), active inflammation, bacterial infection (40–200 mg/L), severe bacterial
infections and burns (>200 mg/L).[26]
CRP is a more sensitive and accurate reflection
of the acute phase response than the ESR (Erythrocyte Sedimentation Rate). The
half-life of CRP is constant. Therefore, CRP level is mainly determined by the
rate of production (and hence the severity of the precipitating cause). In the
first 24 h, ESR may be normal and CRP elevated. CRP returns to normal more
quickly than ESR in response to therapy.
Cardiology diagnostic test
Arterial damage results from white blood cell
invasion and inflammation within the wall. CRP is a general marker for
inflammation and infection, so it can be used as a very rough proxy for heart
disease risk. Since many things can cause elevated CRP, this is not a very
specific prognostic indicator.[27] Nevertheless, a level above 2.4 mg/L has
been associated with a doubled risk of a coronary event compared to levels below
1 mg/L;[2] however, the study group in this case consisted of patients who had
been diagnosed with unstable angina pectoris; whether elevated CRP has any
predictive value of acute coronary events in the general population of all age
ranges remains unclear.
Crioglobulin - the protein of the g-globulin
fraction. Like to the C-reactive protein crioglobulin absent in blood plasma of
the healthy people and occurs at leukoses, rheumatic disease, liver cirrhosis,
nephroses. The characteristic physico-chemical feature of crioglobulin is its
dissolubility at standard body temperature (37 oC) and capacity to form the
sediment at cooling of a blood plasma up to 4 oC.
a2-macroglobulin - protein of a2-globulin
fraction, universal serum proteinase inhibitor. Its contents (2,5 g/l) in blood
plasma is highest comparing to another proteinase inhibitors.
The biological role of a2-macroglobulin consists
in regulation of the tissue proteolysis systems which are very important in
such physiological and pathological processes as blood clotting, fibrinolysis,
processes of immunodefence, functionality of a complement system, inflammation,
regulation of vascular tone (kinine and renin-angiothensine system).
a1-antitrypsin (a1-globulin) – glycoprotein with
a molecular weight 55 kDa. Its concentration in blood plasma is 2-3 г/л. The
main biological property of this inhibitor is its capacity to form complexes
with proteinases oppressing proteolitic activity of such enzymes as trypsin,
chemotrypsin, plasmin, trombin. The content of a1-antitrypsin is markedly
increased in inflammatory processes. The inhibitory activity of a1-antitrypsin
is very important in pancreas necrosis and acute pancreatitis because in these
conditions the proteinase level in blood and tissues is sharply increased. The
congenital deficiency of a1-antitrypsin results in the lung emphysema.
Fibronectin – glycoprotein of blood plasma that
is synthesized and secreted in intercellular space by different cells.
Fibronectin present on a surface of cells, on the basalmembranes, in connective
tissue and in blood. Fibronectin has properties of a «sticking» protein and
contacts with the carbohydrate groups of gangliosides on a surface of plasma
membranes executing the integrative function in intercellular interplay.
Fibronectin also plays important role in the formation of the pericellular
matrix.
Haptoglobin - protein of a2-globulin fraction
of blood plasma. Haptoglobin has
capacity to bind a free haemoglobin forming a complex that refer to b-globulins
electrophoretic fraction. Normal concentration in blood plasma - 0,10-0,35 g/l.
Haptoglobin-hemoglobin complexes are absorbed by
the cells of reticulo-endothelial system, in particular in a liver, and
oxidized to cholic pigments. Such haptoglobinfunction promotes the preservation
of iron ions in an organism under conditions of a physiological and
pathological erythrocytolysis.
Transferrin - glycoprotein belonging to the
b-globulin fraction. It binds in a blood plasma iron ions (Fe3+). The protein
has on the surface two centers of linkage of iron.Transferrin is a transport
form of iron delivering its to places of
accumulation and usage.
Ceruloplasmin - glycoprotein of the a2-globulin
fraction. It can bind the copper ions in blood plasma. Up to 3 % of all copper
contents in an organism and more than 90 % copper contents in plasma is
included in ceruloplasmin. Ceruloplasmin has properties of ferroxidase
oxidizing the iron ions. The decrease of
ceruloplasmin in organism (Wilson disease) results in exit of copper ions from
vessels and its accumulation in the connective tissue that shows by
pathological changes in a liver, main brain, cornea.
The place of synthesis of each fraction and
subfruction of blood plasma proteins.
Albumins, a1-globulins, fibrinogen are fully
synthesized in hepatocytes. Immunoglobulins are produced by plasmocytes (immune
cells). In liver cryoglobulins and some other
g-globulins are produced too.
a2-globulins and b-globulins are partly synthesized in liver and partly
in reticuloendothelial cells.
Causes and consequences of protein content
changes in blood plasma.
Hypoproteinemia
- decrease of the total contents of proteins in blood plasma. This state
occurs in old people as well as in pathological states accompanying with the
oppressing of protein synthesis (liver diseases) and activation of
decomposition of tissue proteins (starvation, hard infectious diseases, state
after hard trauma and operations, cancer). Hypoproteinemia (hypoalbuminemia)
also occurs in kidney diseases, when the increased excretion of proteins via
the urine takes place.
Hyperproteinemia
- increase of the total contents of proteins in blood plasma. There are
two types of hyperproteinemia - absolute
and relative.
Absolute hyperproteinemia – accumulation of the
proteins in blood. It occurs in infection and inflammatory diseases
(hyperproduction of immunoglobulins),
rheumatic diseases (hyperproduction of C-reactive protein), some
malignant tumors (myeloma) and others.
Relative hyperproteinemia – the increase of the
protein concentration but not the absolute amount of proteins. It occurs when
organism loses water (diarrhea, vomiting, fever, intensive physical activity
etc.).
The principle of the measurement of protein
fractions by electrophoresis method.
Electrophoresis is the separation of proteins on
the basis of their electric charge. It depends ultimately on their base-acid
properties, which are largely determined by the number and types of ionizable R
groups in their polipeptide chains. Since proteins differ in amino acid
composition and sequence, each protein has distinctive acid-base properties.
There are a number of different forms of electroforesis useful for analyzing
and separating mixtures of proteins
If a precursor of an antibody-secreting cell
becomes cancerous, it divides uncontrollably to generate a clone of plasma
cellssecreting a single kind of antibody molecule. The image (courtesy of
Beckman Instruments, Inc.) shows — from left to right — the electrophoretic
separation of:
1.
normal human serum with its diffuse band of gamma globulins;
2.
serum from a patient with multiple myeloma producing an IgG myeloma
protein;
3.
serum from a patient with Waldenström's macroglobulinemia where the
cancerous clone secretes an IgM antibody;
4.
serum with an IgA myeloma protein.
§
Gamma globulins can be harvested from donated blood (usually pooled from
several thousand donors) and injected into persons exposed to certain diseases
such as chicken pox and hepatitis. Because such preparations of immune globulin
contain antibodies against most common infectious diseases, the patient gains
temporary protection against the disease.
Serum Lipids
Because of their relationship to cardiovascular
disease, the analysis of serum lipids has become an important health measure.
The table shows the range of typical values as
well as the values above (or below) which the subject may be at increased risk
of developing atherosclerosis.
•
Total cholesterol is the serum of blood
o HDL
cholesterol
o LDL
cholesterol and
o 20%
of the triglyceride value
•
Note that
o
high LDL values are bad, but
o
high HDL values are good.
•
Using the various values, one can calculate a
cardiac risk ratio = total cholesterol divided by
HDL cholesterol
A cardiac risk ratio greater than 7 is considered
a warning
A. Protein fractions which are
received by the electrophoresis
B. Protein fractions which are received with the help of
imunoelectropheresis on agar
gel.
Residual nitrogen, its components, ways of their
formation, blood content
The state of protein nutrition can be determined
by measuring the dietary intake and output of nitrogenous compounds from the
body. Although nucleic acids also contain nitrogen, protein is the major
dietary source of nitrogen and measurement of total nitrogen intake gives a
good estimate of protein intake (mg N Ч 6.25 = mg protein, as nitrogen is 16%
of most proteins). The output of nitrogen from the body is mainly in urea and
smaller quantities of other compounds in urine and undigested protein in feces,
and significant amounts may also be lost in sweat and shed skin.
The difference between intake and output of
nitrogenous compounds is known as nitrogen balance. Three states can be
defined: In a healthy adult, nitrogen balance is inequilibrium when intake
equals output, and there is no change in the total body content of protein. In
a growing child, a pregnant woman, or in recovery from protein loss, the
excretion of nitrogenous compounds is less than the dietary intake and there is
net retention of nitrogen in the body as protein, ie, positive nitrogen
balance. In response to trauma or infection or if the intake of protein is
inadequate to meet requirements there is net loss of protein nitrogen from the
body, ie, negative nitrogen balance. The continual catabolism of tissue
proteins creates the requirement for dietary protein even in an adult who is
not growing, though some of the amino acids released can be reutilized.
Nitrogen balance studies show that the average
daily requirement is
Residual nitrogen – nonprotein nitrogen, that is
nitrogen of organic and inorganic compounds that remain in blood after protein
sedimentation.
Organic and inorganic compounds of residual
nitrogen are as follows: urea (50 % of the residual nitrogen), amino acids (25
%), creatine and creatinine (7,5 %), salts of ammonia and indicane (0,5 %),
other compounds (about 13 %).
Urea is formed in liver during the degradation of
amino acids, pyrimidine nucleotides and other nitrogen containing compounds.
Amino acids are formed as result of protein decomposition or owing to the
conversion of fatty acids or carbohydrates to amino acids. The pool of amino
acids in blood is also supported by the process of their absorption in
intestine. Creatine is produced in kidneys and liver from amino acids glycine
and arginine, creatinine is formed in muscles as result of creatine phosphate
splitting. In result of ammonia neutralization the ammonia salts can be formed.
Indicane is the product of indol neutralization in the liver.

Creatinine
Urine
The content of
residual nitrogen in blood is 0,2 – 0,4 g/l.
The pathways of convertion of amino acid
nonnitrogen residues.
The removal
of the amino group of an amino acid by transamination or oxidative deamination
produces an α-keto acid that contains the carbon skeleton from the amino
acid (nonnitrogen residues). These α-keto acids can be used for the
biosynthesis of non-essential amino acids or undergoes a different degradation
process. For alanine and serine, the degradation requires a single step. For
most carbon arrangements, however, multistep reaction sequences are
required. There are only seven
degradation sequences for 20 amino acids. The seven degradation products are
pyruvate, acetyl CoA, acetoacetyl CoA, α-ketoglutarate, succinyl CoA,
fumarate, and oxaloacetate. The last four products are intermediates in the
citric acid cycle. Some amino acids have more than one pathway for degradation.
The major
point of entry into the tricarboxylate cycle is via acetyl-CoA; 10 amino acids
enter by this route. Of these, six (alanine, glycine, serine, threonine,
tryptophan and cysteine) are degraded to acetyl-CoA via pyruvate, five
(phenylalanine, tyrosine, leucine, lysine, and tryptophan) are degraded via
acetoacetyl-CoA, and three (isoleucine, leucine and tryptophan) yield
acetyl-CoA directly. Leucine and
tryptophan yield both acetoacetyl-CoA and acetyl-CoA as end products.
The carbon
skeletons of five amino acids (arginine, histidine, glutamate, glutamine and
proline) enter the tricarboxylic acid cycle via a-ketoglutarate.
The carbon
skeletons of methionine, isoleucine, and valine are ultimately degraded via
propionyl-CoA and methyl-malonyl-CoA to succinyl-CoA; these amino acids are
thus glycogenic.
Fumarate is
formed in catabolism of phenylalanine, aspartate and tyrosine.
Oxaloacetate
is formed in catabolism of aspartate and asparagine. Aspartate is converted to
the oxaloacetate by transamination.
Amino acids
that are degraded to citric acid cycle intermediates can serve as glucose
precursors and are called glucogenic. A glucogenic amino acid is an amino acid
whose carbon-containing degradation product(s) can be used to produce glucose
via gluconeogenesis.
Amino acids
that are degraded to acetyl CoA or acetoacetyl CoA can contribute to the
formation of fatty acids or ketone bodies and are called ketogenic. Aketogenic
amino acid is an amino acid whose carbon-containing degradation product(s) can
be used to produce ketone bodies.
Amino acids
that are degraded to pyruvate can be either glucogenic or ketogenic. Pyruvate
can be metabolized to either oxaloacetate (glucogenic) or acetyl CoA
(ketogenic).
Only two
amino acids are purely ketogenic: leucine and lysine. Nine amino acids are both
glucogenic and ketogenic: those degraded to pyruvate (alanine, glycine,
cysteine, serine, threonine, tryptophan), as well as tyrosine, phenylalanine,
and isoleucine (which have two degradation products). The remaining nine amino
acids are purely glucogenic (arginine, asparagine, aspartate, glutamine,
glutamate, valine, histidine,
methionine, proline)
The
regulation of protein metabolism. Protein metabolism is regulated by different
hormones. All hormones according to their action on protein synthesis or
splitting are divided on two groups: anabolic and catabolic. Anabolic hormones
promote to the protein synthesis. Catabolic hormones enhance the decomposition
of proteins.
Somatotropic
hormone (STH, growth hormone):
- stimulates the passing of amino
acids into the cells;
- activates the synthesis of proteins, DNA,
RNA.
Thyroxine and
triiodthyronine:
- in normal concentration stimulate
the synthesis of proteins and nucleic acids;
- in excessive concentration
activate the catabolic processes.
Insulin:
- increases the permeability of cell
membranes for amino acids;
- activates synthesis of proteins
and nucleic acids;
- inhibits the conversion of amino
acids into carbohydrates.
Glucagon:
- stimulates the conversion of amino
acids into carbohydrates.
Epinephrine:
- activates
the protein decomposition.
Glucocorticoids:
- stimulate the catabolic processes
(protein decomposition) in connective, lymphoid and muscle tissues and activate
the processes of protein synthesis in liver;
- stimulate the activity of
aminotransferases;
- activate the synthesis of urea.
Sex hormones:
- stimulate the processes of
protein, DNA, RNA synthesis;
- cause the positive nitrogenous
balance.
The role of
liver in protein metabolism:
– synthesis of plasma
proteins. Most of plasma proteins are
synthesized in liver: all albumins, 75-90 % of α-globulins, 50 % of
β-globulins, all proteins of blood clotting systems (prothrombin,
fibrinogen, proconvertin, proaccelerine). Only γ-globulins are synthesized
in the cells of reticuloendothelial system.
– synthesis of urea and
uric acid;
– synthesis of choline and
creatine;
– transamination and
deamination of amino acids.
Clinical
significance of residual nitrogen measurement in blood. The kinds of azotemia.
Azotemia - increase of the residual nitrogen content
in blood. There are two kinds of azotemia: absolute and relative.
Absolute
azotemia – accumulation of the components of residual nitrogen in blood.
Relative azotemia occurs in dehydration of the organism (diarrhea, vomiting).
Absolute
azotemia can be divided on the productive azotemia and retention azotemia.
Retention azotemia is caused by the poor excretion of the nitrogen containing
compounds via the kidneys; in this case the entry of nitrogen containing
compounds into the blood is normal.
Retention
azotemia can be divided on the renal and extrarenal. Renal retention azotemia
occurs in kidney diseases (glomerulonephritis, pyelonephritis, kidney
tuberculosis et c.). Extrarenal retention azotemia is caused by the violations
of kidney hemodynamic and decrease of glomerulus filtration processes (heart
failure, local disorders of kidney hemodynamic).
Productive
azotemia is conditioned by the enhanced entry of nitrogen containing compounds
into the blood. The function of kidneys in this case doesn’t suffer.Productive
azotemia can be observed in cachexia, leukoses, malignant tumors, treatment by
glucocorticoids.
Prerenal
Azotemia
Alternate
Names : Azotemia - Prerenal, Renal Underperfusion, Uremia
|
|

{kind=link}
{kind=link}
![All peripheral cells express the LDL-receptor (LDLR), and recycle it to the cell surface upon need for cholesterol. Cholesterol is delivered to these cells through binding of LDL to LDLR, which triggers endocytosis (internalization) of both species. When the need for cholesterol is satisfied, the recycling of LDLR is discontinued. Normally, an LDL particle stays in circulation for no more than a few days before being consumed by a cholesterol needing cell. However, under conditions of sustained cholesterol excess, the particle stays in circulation for longer periods of time, and becomes more vulnerable to undesired modifications (e.g. oxidation). As high levels of oxidized LDL are commonly found in atherosclerotic plaques, they are thought to be the major inducer of atherosclerotic lesions. Hence, LDL became known as bad cholesterol. However, today we know that not all LDL particles are bad, and that some LDL particles, especially very large ones (with diameter >21.3nm), may even provide protection against atherosclerosis (2). LDL and HDL particle sizes are largely determined by a LP-associated protein, called CETP (cholesteryl ester transfer protein). This protein enhances exchange of non-polar lipids, primarily CE and TG, and facilitates tight packaging of CE within the core of the particles. The end result of prolonged and/or efficient CETP action is smaller LDL and HDL particles. [The LP-anchored CETP can be envisioned as having a hand that rotates between the interior and exterior of the particle and capable of holding only one lipid molecule at a time. Grasping of one molecule releases another and vise versa.]](http://intranet.tdmu.edu.ua/data/kafedra/internal/chemistry/classes_stud/en/med/lik/ptn/2/Investigation%20of%20acid-base%20state%20of%20blood%20and%20respiratory%20function%20of%20erythrocytes..files/image039.jpg){kind=link}
{kind=link}

http://www.youtube.com/watch?v=97uiV4RiSAY
http://www.youtube.com/watch?v=-WhADd1GKtA&feature=relmfu
{kind=link}



Although
These Researchers Beg to Differ…
{kind=link}
{kind=link}
http://www.youtube.com/watch?v=XLLBlBiboJI&feature=related
http://www.youtube.com/watch?v=-WhADd1GKtA&feature=relmfu
Here
is a diagram of Low-Density Lipoprotein (LDL) which is approximately 25nm in
diameter:
http://www.youtube.com/watch?v=x-4ZQaiZry8

You
need "Quick Time" Player and
Plug-In to
view this LDL particle in motion:
http://www.youtube.com/watch?v=97uiV4RiSAY
{kind=link}
Characteristics
of Lipoproteins in Human Plasma
{kind=link}
Biochemistry of immune processes.
http://www.youtube.com/watch?v=Ys_V6FcYD5I&feature=related
http://www.youtube.com/watch?v=14koX2tbRzU&feature=related
http://www.youtube.com/watch?v=VOD5tuQ5wvo&feature=related





![Factor C3, the products of which are involved in several functions, is central to the complement system. The classic pathway is triggered by the formation of factor C1 at IgG or IgM on the surface of microorganisms (left). C1 is an 18-part molecular complex with three different components (C1q, C1r, and C1s). C1q is shaped like a bunch of tulips, the “flowers” of which bind to the Fc region of antibodies (left). This activates C1r, a serine proteinase that initiates the cascade of the classic pathway. First, C4 is proteolytically activated into C4b, which in turn cleaves C2 into C2a and C2b. C4B and C2a together form C3 convertase [1], which finally catalyzes the cleavage of C3 into C3a and C3b. Small amounts of C3b also arise from non-enzymatic hydrolysis of C3.](http://intranet.tdmu.edu.ua/data/kafedra/internal/chemistry/classes_stud/en/med/lik/ptn/2/Investigation%20of%20acid-base%20state%20of%20blood%20and%20respiratory%20function%20of%20erythrocytes..files/image069.jpg){kind=link}
http://www.youtube.com/watch?v=lrYlZJiuf18
http://www.youtube.com/watch?v=Ys_V6FcYD5I&feature=related

{kind=link}
http://www.youtube.com/watch?v=mUXIK5gGD1k


Biosynthesis of a light chain _
Training
the immune system -- the "J" curve

Stress
is cumulative

Recommendations
{kind=link}
· Heed your body's early warning signs,
o Disordered sleep (too much or
insomnia)
o Loss of interest in pleasurable
activities
o Poor concentration. Lack of mental
energy.
· Get an annual influenza vaccine
(usually available each year starting in October)