Anaerobic and aerobic oxidation of glucose.
Alternative
ways of monosaccharides metabolism.
Foods high in
carbohydrate include fruits, sweets, soft drinks, breads, pastas, beans,
potatoes, bran, rice, and cereals. Carbohydrates are a common source of energy
in living organisms; however, no carbohydrate is an essential nutrient in humans.
Carbohydrates
are not necessary building blocks of other molecules, and the body can obtain
all its energy from protein and fats.[ The brain and neurons generally
cannot burn fat for energy, but use glucose or ketones.
Humans can synthesize some glucose (in a set of processes known as gluconeogenesis)
from specific amino acids, from the glycerol backbone
in triglycerides and
in some cases from fatty acids. Carbohydrate and protein contain 4 calories
per gram,
while fats contain 9 calories per gram. In the case of protein, this is
somewhat misleading as only some amino acids are usable for fuel.
Organisms
typically cannot metabolize all types of carbohydrate to yield energy. Glucose
is a nearly universal and accessible source of calories.
Many organisms also have the ability to metabolize other monosaccharides and Disaccharides,
though glucose is preferred. In Escherichia coli,
for example, the lac operon will express enzymes for the digestion of
lactose when it is present, but if both lactose and glucose are present
the lac operon is repressed, resulting in the glucose being used
first. Polysaccharides are also common sources of energy. Many
organisms can easily break down starches into glucose, however, most organisms
cannot metabolize cellulose or other polysaccharides like chitin and arabinoxylans.
These carbohydrates types can be metabolized by some bacteria and
protists. Ruminants and termites,
for example, use microorganisms to processcellulose
Even though these complex
carbohydrates are not very digestible, they represent an important dietary
element for humans, called dietary fiber. Fiber enhances digestion, among other
benefits.
Based on the
effects on risk of heart disease and obesity, the Institute of Medicine recommends that
American and Canadian adults get between 45–65% of dietary energy from
carbohydrates. The Food and Agriculture Organization and World Health Organization jointly
recommend that national dietary guidelines set a goal of 55–75% of total energy
from carbohydrates, but only 10% directly from sugars (their term for simple
carbohydrates).
The biological significance of carbohydrates in
living organisms
 A carbohydrate is an organic compound that is composed of
atoms of carbon, hydrogen and oxygen in a ratio of 1 carbon atom, 2 hydrogen
atoms, and 1 oxygen atom. Some carbohydrates are relatively small molecules,
the most important to us is glucose which has 6 carbon atoms. These simple
sugars are called monosaccharides.
A carbohydrate is an organic compound that is composed of
atoms of carbon, hydrogen and oxygen in a ratio of 1 carbon atom, 2 hydrogen
atoms, and 1 oxygen atom. Some carbohydrates are relatively small molecules,
the most important to us is glucose which has 6 carbon atoms. These simple
sugars are called monosaccharides.
The primary function of carbohydrates is for
short-term energy storage
(sugars are for Energy). A secondary function is intermediate-term energy
storage (as in starch for plants and glycogen for animals). Other carbohydrates
are involved as structural components in cells, such as cellulose which is
found in the cell walls of plants.
Two common Monosaccharides, (single sugars) Glucose
and Fructose
Hooking two monosaccharides together forms a more complex
sugar, such as the union of glucose and fructose to give sucrose, or common
table sugar. Compounds such as sucrose are called Disaccharides (two
sugars). Both monosaccharides and disaccharides are soluble in water.
Larger, more complex carbohydrates are formed
by linking shorter units together to form long or very long sugar chains called
Polysaccharides. Because of their size, these are often times not
soluble in water. Many biologically important compounds such as starches and cellulose are Polysaccharides. Starches
are used by plants, and glycogen by animals, to store energy in their numerous
carbon-hydrogen bonds, while cellulose is an important compound that adds
strength and stiffness to a plant's cell wall.
Sugars are most often found in the form of a
"RING". The glucose molecule in the image above and the one in the
image below (Glc) are really the same molecule, just arranged differently. The
corners of the "stop sign" represent Carbon atoms even thought they
are not labeled with a "C" (its chemistry shorthand). To form
these rings, the Carbonyl (C=0) Carbon of the straight-chain form (above) forms a bond
with the next to last Carbon in the chain, making the ring.
The image on the left shows two monosaccharides,
Glucose and Galactose (Gal). Examine their structure and you will notice there
is very little difference. Their molecular formulas, C6H1206, are even the
same. Molecules
with the same chemical formula, but different molecular structures
are called Isomers.
The sugar subunits can be linked by the reaction,
dehydration synthesis, to form larger molecules. The disaccharide, Sucrose, is
formed from two monosaccharides, Glucose and Fructose.
The disaccharide Lactose is a dimer (two subunits) of Glucose and Galactose, the disaccharide Maltose is a dimer of
Glucose.
Large polymers of sugars are called Carbohydrates.
Carbohydrates can be 100's of sugars long and either straight or branched. The
term Complex Carbohydrate, or sometimes even just Carbohydrate refers to
long chains of sugars. Three common types of complex carbo's we will examine
are: Starch, Cellulose, and Glycogen. All three are composed only of Glucose.
They differ only in the bonding arrangements between the Glucose subunits. Not
all complex carbs are composed of glucose alone, many have highly unusual
sugars in their chains.
Starch is a long
(100's) polymer of Glucose molecules, where all the sugars are oriented in the
same direction. Starch is one of the primary sources of calories for humans.
Cellulose is a long (100's) polymer of Glucose molecules. However the orientation
of the sugars is a little different. In Cellulose, every other sugar molecule
is "upside-down". This small difference in structure makes a big
difference in the way we use this molecule.
Glycogen is another Glucose polymer. Glycogen is a stored energy source, found in
the Liver and muscles of Humans. Glycogen is different from both Starch and
Cellulose in that the Glucose chain is branched or "forked".
As we noted, one function of
carbohydrates (such as sugars) is for Energy. A secondary function is
intermediate-term energy storage (as in starch for plants and
glycogen for animals). Often the energy content of sugars is used to justify
downing a Snickers Bar (or two). On the other hand, complex carbs that must be
broken down before the sugars can be used are thought of as "Slow"
Energy. The simple sugars are gradually released over time, providing a slow but
steady source of Energy.
Pastas and
whole-grain breads contain complex carbohydrates, which are long strands of
glucose molecules.
Nutritionists
recommend that 55–60 percent of calories come from carbohydrates, and
especially complex carbohydrates.
All animals derive the major portion of
their food calories from the different types of Carbohydrates in their diets.
Most of the energy for the metabolic
activities of the cell in all organisms is derived from the oxidation of
Carbohydrate. Important functions of Carbohydrate are that of storing food,
acting as a framework in body, performs are listed below.
Carbohydrate
functions as Bio Fuel
Carbohydrate
functions as an energy source of the body and acts as Bio fuel.Step wise
details for the process of production of energy are discussed below.
·
Polysaccharides such as starch
and glycogen are first hydrolyzed by enzymes to Glucose.
·
Glucose is the transported from
one cell to another by blood in case of animals and cell sap in case of plants.
·
Glucose is then oxidized to
produce carbon dioxide and water.
·
Energy is released in this
process which is used for functioning of the cells.
The process
of production of energy by carbohydrates is described in above steps. Now it is
important to note, that fats and proteins can also be burned to provide energy
but carbohydrate functions as primary
source of energy. Fats are only burned if there is non availability
of carbohydrates. When fat is burned in absence of carbohydrates, toxic
compounds like called ketone bodies are
produced. Accumulation of these ketone bodies over long period causes a
condition called Ketosis. In this condition blood becomes unable to
carry oxygen properly and this can be fatal. Thus, one of important function of
carbohydrate is help burn fat properly.
Different
forms of Carbohydrate are stored in living organism as storage food.
·
Polysaccharide starch acts as storage
food for plants.
·
Glycogen stored in liver and
muscles acts as storage food for animals.
·
Inulin acts as storage food of
dahlias, onion and garlic.
Thus
carbohydrate performs the function of storing food.
Different
Carbohydrates especially Polysaccharides act as framework in living organism.
·
Cellulose forms cell wall of
plant cell along with hemicelluloses and Pectin
·
Chitin forms cell
wall of fungal cell and exoskeleton of arthropods
·
Peptidoglycan forms cell wall
of bacteria and cyanobacteria.
Thus
carbohydrates function as contributing material to the cellular structure.
Heparin is a
polysaccharide (carbohydrate) which acts as anticoagulant and prevents
intravascular clotting.
Many antigens
are glycoprotein (which contains oligosaccharide) in nature and give
immunological properties to the blood.
Many Hormones
like FSH (Follicular Stimulating Hormone which takes part in ovulation in
females) and LH (Leutinizing Hormone) are glycoprotein and help in reproductive
processes.
Carbohydrates
are an important component of many industries like textile, paper, lacquers and
breweries.
Agar is polysaccharide used in culture media, laxative and food.
Cellulose acts as roughage of food. It stimulates peristalsis movement and
secretion of digestive enzymes.
Hyaluronic
acid found in between joints acts as synovial fluid and provides frictionless
movement.
Carbohydrates
have the general molecular formula CH2O, and thus were once thought
to represent "hydrated carbon". However, the arrangement of atoms in
carbohydrates has little to do with water molecules.
http://www.youtube.com/watch?v=p-lFJVOkFwk
Digestion of carbohydrates: localization,
types, role of enzymes.
In simple terms, our digestion system - from the mouth to the small
intestine - is designed to break down disaccharides and polysaccharides into
monosaccharides. This metabolism of carbohydrates is achieved through the
secretion of a number of digestive enzymes into the gastrointestinal
tract (especially in the duodenum) where they attack carbohydrates and
gradually convert them into simple sugars like glucose so they can be
absorbed into the blood. Digestive enzymes are like biological scissors - they
chop long starch molecules into simpler ones.
In the Mouth
The process of
digesting carbohydrates begins in the mouth. Our saliva contains an enzyme
called amylase that starts breaking down the more complex carbs into
simpler types.
In the Stomach
Enzyme activity continues in the stomach, but slows down significantly as digestive acids
are released into the stomach by the glands.
In the Small
Intestine
Another version of amylase is secreted by the pancreas
into the duodenum (first section of small intestine). This cuts down
carbohydrates into simple sugars - maltose, lactose
and sucrose.
As the carbohydrate passes further into the intestine, the enzymes maltase,
lactase
and sucrase
chop maltose, lactose and sucrose into smaller bits, more easily absorbed,
which are eventually converted to glucose and absorbed through the intestinal
walls into the bloodstream.
Glucose Metabolism
By The Liver
After
carbohydrates are duly broken down into glucose, in the duodenum and jejunum of
the small intestine, the glucose is absorbed into the bloodstream and taken to
the liver, where it is stored or distributed to cells throughout the body for
energy. In this way, the liver regulates blood glucose levels to provide
sufficient energy for the body. For example, excess glucose (a cause of
hyperglycemia) is converted in the liver to glycogen (glycogenolysis) in
response to the hormone insulin, and stored. Likewise, if blood sugar
levels fall, (eg. between meals), the glycogen is re-converted to glucose
(glycogenolysis) in response to messages conveyed by the hormone glucagon,
to prevent hypoglycemia. If glycogen levels are exhausted, glucagon can trigger
the formation of glucose from some amino acids (protein) or glycerol (fats) - a
process called gluconeogenesis.
The primary
organ responsible for the regulation of blood glucose levels is the liver.
Blood glucose levels must be maintained in the range of 80-120 mg/100 ml.
In order to accomplish this the liver is capable of taking up large amounts of
glucose. The enzyme glucokinase is responsible for doing this. Glucokinase has
a high Km for glucose and is not inhibited by the product of the
reaction glucose 6-phosphate so even when the serum glucose level is high,
glucokinase remains active. The glucose 6-phosphate formed in the liver is
trapped there because the phosphorylated derivative cannot cross the plasma
membrane. In addition, the enzyme phosphorylase a acts as the glucose sensor for the
body and is allosterically inactivated by high glucose concentrations. The fate
of glucose 6-phosphate is controlled by the levels of insulin and
glucogon. The figure below compares the control of blood glucose levels by the
liver after eating and after an overnight fast.
What Determines
Speed of Carb Digestion
Generally speaking, the speed of
digestion is determined by the chemical nature of the carbohydrate itself, and
thus how "resistant" it is to the activity of the enzymes. A simple
sugar is much less resistant than a starch, and is digested or metabolized much
faster. Things that slow down digestion include: the presence of acid
(from gastric juices or the food itself), and the presence of soluble fiber.
Digestion
of cellulose
Fiber- What is
fiber? What your parents used to call roughage, food companies now promote as
fiber. Fiber is an undigestable complex carbohydrate found in plants.
Fiber is not a single food or substance. Fiber in itself has no calories
because the body cannot absorb it. Therefore, high fiber foods, such as fruits
and vegetables, are low in fat and low in calories. Fiber can be divided into
two categories according to their physical characteristics and effects on the
body: Water insoluble and water soluble. Each form functions differently and
provides different health benefits. Insoluble fibers, such as cellulose,
hemicellulose and lignin, do not dissolve in water. Soluble fibers, such as
gums and pectins, do dissolve in water. Dietary Fiber is composed of undigestable
complex carbohydrates. There are two basic types of fiber. Soluble Fiber - Pectins, acidic sugars often
found in fruits. Insoluble fiber - Cellulose is one example, these are often found in the
body of plants and in seed coats (where they are also known as bran).
There's a lot of
confusion surrounding carbohydrates: Are they good or bad for us? Is a low-carb
diet a good way to lose weight?
You need
carbohydrates to function. They are the body's primary energy source and used
for both physical activity and normal body functions such as brain function,
heartbeat, breathing and digestion.
To eat or not to
eat? There's a lot of confusion surrounding carbohydrates.
There is a huge difference
between the natural "good" carbs that our bodies need to function and
the unnatural, highly-processed, "refined" carbs so many of us eat
every day!
The following food types are considered
good carbohydrates: whole vegetables and fruits, beans, legumes and whole
grains.
They include a
range of the following healthy characteristics:
- High in fibre:
These foods make you feel full, take longer to digest and cause a slow rise in
blood sugar. They also promote waste elimination and help your body get rid of
toxins. Good sources include raw fruits and vegetables with their skins,
legumes, whole grains, berries and dried fruits.
- Low to
moderate glycemic index: Good carbs stabilize blood sugar levels and insulin
production.
- High in
nutrients: They include natural vitamins, minerals and phytonutrients that
promote good health
- Low calorie:
They provide sustained energy, promote healthy weight loss and long-term weight
maintenance
- Greater
thermic-effect: They naturally stimulate metabolism and promote fat loss
BAD
CARBOHYDRATES
Carbohydrates are bad
for your skin
Bad carbs are refined, processed
carbohydrate foods that have had all or most of their natural nutrients and
fibre removed.
In addition to
providing your body with empty calories, eating too many of these refined
carbohydrates causes your blood sugar levels to spike, sending a signal to the
pancreas to over-secrete insulin. In a nutshell, this facilitates the excess
storage of fat.
And that's not
all. A blood-sugar spike caused by refined carbs, followed by an over-secretion
of insulin to combat the spike will result in an energy crash. This ultimately
becomes a vicious cycle as you grab something sugary to try and bring your
blood sugar levels back up. Bleached wheat flour, white sugar, artificial
flavouring and preservatives are the most common ingredients used to make
"bad carb" foods. Examples are the white versions of baked goods,
breads and pastas, as well as snack foods, sugary cereals and soft drinks.
ESTIMATING HOW
MANY CARBOHYDRATE CALORIES YOU SHOULD EAT
Carbohydrates contain
four calories per gram. If you eat a 2,000-calorie-per-day diet and are aiming
to eat 55 per cent of your calories from carbs, plan for 1,100 calories to come
from carbs (275 grams).
Opt for whole
grain versions of bread products, rice, pasta and cereals. Skip white sugar all
together.
The mechanism of monosaccharides absorption
Simple sugars are far and away the
predominant carbohydrate absorbed in the digestive tract, and in many animals
the most important source of energy. Monosaccharides, however, are only rarely
found in normal diets. Rather, they are derived by enzymatic digestion of more
complex carbohydrates within the digestive tube.
Particularly
important dietary carbohydrates include starch and disaccharides such as
lactose and sucrose. None of these molecules can be
absorbed for the simple reason that they cannot cross cell membranes unaided
and, unlike the situation for monosaccharides, there are no transporters to
carry them across.
This section
will focus on understanding the processes involved in assimilation of three
important carbohydrates: starch, lactose and sucrose. The key concepts involved
in all three cases are that:
Polysaccharides and disaccharides must
be digested to monosaccharides prior to absorption and the key players in these
processes are the brush border hydrolases, which include maltase, lactase and
sucrase. Dietary lactose and sucrose are "ready" for digestion by
their respective brush border enzymes. Starch, as discussed previously, is
first digested to maltose by amylase in pancreatic secretions and, in some
species, saliva.
Dietary
lactose and sucrose, and maltose derived from digestion of starch, diffuse in
the small intestinal lumen and come in contact with the surface of absorptive
epithelial cells covering the villi where they engage with brush border
hydrolases:
At long last,
we're ready to actually absorb these monosaccharides. Glucose and galactose are
taken into the enterocyte by cotransport with sodium using the same
transporter. Fructose enters the cell from the intestinal lumen via facilitated
diffusion through another transporter.
Absorption of glucose entails transport
from the intestinal lumen, across the epithelium and into blood. The
transporter that carries glucose and galactose into the enterocyte is the
sodium-dependent hexose transporter, known more formally as SGLUT-1. As the name
indicates, this molecule transports both glucose and sodium ion into the cell
and in fact, will not transport either alone.
The essence
of transport by the sodium-dependent hexose transporter involves a series of
conformational changes induced by binding and release of sodium and glucose,
and can be summarized as follows:
1. the
transporter is initially oriented facing into the lumen - at this point it is
capable of binding sodium, but not glucose
2. sodium binds,
inducing a conformational change that opens the glucose-binding pocket
3. glucose binds
and the transporter reorients in the membrane such that the pockets holding
sodium and glucose are moved inside the cell
4. sodium dissociates
into the cytoplasm, causing glucose binding to destabilize
5. glucose
dissociates into the cytoplasm and the unloaded transporter reorients back to
its original, outward-facing position
The animation
seen below depicts digestion of maltose and entry of the resulting glucose,
along with sodium, into the enterocyte (actually, two sodium ions are
transported for each glucose). Despite the simplicity of the diagram, you
should easily be able to identify the sodium-dependent hexose transporter and
"watch" its conformational changes. Also, imagine the corresponding
process involving lactose and sucrose assimilation.
Fructose is not co-transported with
sodium. Rather it enters the enterocyte by another hexose transporter (GLUT5).
Once inside
the enterocyte, glucose and sodium must be exported from the cell into blood.
We've seen previously how sodium is rapidly shuttled out in exchange for
potassium by the battery of sodium pumps on the basolateral membrane, and how
that process maintains the electrochemical gradient across the
epithelium. The energy stored in this gradient is actually what
is driving glucose entry through the sodium-dependent hexose transporter
described above. Recall also how the massive transport of sodium out of the
cell establishes the osmotic gradient responsible for absorption of water.
Glucose,
galactose and fructose are tranported out of the enterocyte through another
hexose transporter (called GLUT-2) in the basolateral membrane. These
monosaccharides then diffuse "down" a concentration gradient into
capillary blood within the villus.
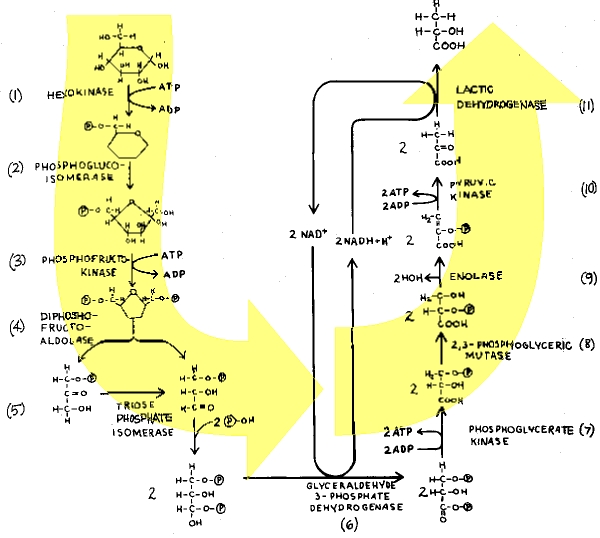
Decomposition of glucose in anaerobic conditions (glycolysis):
The Energy Derived from Glucose Oxidation
Aerobic glycolysis
of glucose to pyruvate, requires two equivalents of ATP to activate the
process, with the subsequent production of four equivalents of ATP and two
equivalents of NADH. Thus, conversion of one mole of glucose to two moles of
pyruvate is accompanied by the net production of two moles each of ATP and
NADH.
Glucose + 2
ADP + 2 NAD+ + 2 Pi -----> 2 Pyruvate + 2 ATP + 2 NADH
+ 2 H+
The NADH
generated during glycolysis is used to fuel mitochondrial ATP synthesis via oxidative
phosphorylation, producing either two or three equivalents of ATP
depending upon whether the glycerol
phosphate shuttle or the malate-aspartate
shuttle is used to transport the electrons from cytoplasmic NADH into
the mitochondria. The net yield from the oxidation of 1 mole of glucose to 2
moles of pyruvate is, therefore, either 6 or 8 moles of ATP. Complete oxidation
of the 2 moles of pyruvate, through the TCA
cycle, yeilds an additional 30 moles of ATP; the total yield, therefore
being either 36 or 38 moles of ATP from the complete oxidation of 1 mole of
glucose to CO2 and H2O.
back
to the top
The
Individual Reactions of Glycolysis
The pathway
of glycolysis can be seen as consisting of 2 separate phases. The first is the
chemical priming phase requiring energy in the form of ATP, and the second is
considered the energy-yielding phase. In the first phase, 2 equivalents of ATP
are used to convert glucose to fructose 1,6-bisphosphate (F1,6BP). In the
second phase F1,6BP is degraded to pyruvate, with the production of 4 equivalents
of ATP and 2 equivalents of NADH.
http://www.youtube.com/watch?v=6JGXayUyNVw&feature=related
http://www.youtube.com/watch?v=nKgUBsC4Oyo&feature=related
Pathway of
glycolysis from glucose to pyruvate. Substrates and products are in blue,
enzymes are in green. The two high energy intermediates whose oxidations are
coupled to ATP synthesis are shown in red (1,3-bisphosphoglycerate and
phosphoenolpyruvate).
http://www.youtube.com/watch?v=PQMsJSme780&feature=related
The
Hexokinase Reaction:
The
ATP-dependent phosphorylation of glucose to form glucose 6-phosphate (G6P)is
the first reaction of glycolysis, and is catalyzed by tissue-specific
isoenzymes known as hexokinases. The phosphorylation accomplishes two goals:
First, the hexokinase reaction converts nonionic glucose into an anion that is
trapped in the cell, since cells lack transport systems for phosphorylated
sugars. Second, the otherwise biologically inert glucose becomes activated into
a labile form capable of being further metabolized.
Four
mammalian isozymes of hexokinase are known (Types I - IV), with the Type IV
isozyme often referred to as glucokinase. Glucokinase is the form of the enzyme
found in hepatocytes. The high Km of glucokinase for glucose means
that this enzyme is saturated only at very high concentrations of substrate.
Comparison of
the activities of hexokinase and glucokinase. The Km for hexokinase
is significantly lower (0.1mM) than that of glucokinase (10mM). This difference
ensures that non-hepatic tissues (which contain hexokinase) rapidly and
efficiently trap blood glucose within their cells by converting it to
glucose-6-phosphate. One major function of the liver is to deliver glucose to
the blood and this in ensured by having a glucose phosphorylating enzyme
(glucokinase) whose Km for glucose is sufficiently higher that the
normal circulating concentration of glucose (5mM).
This feature
of hepatic glucokinase allows the liver to buffer blood glucose. After meals, when postprandial blood glucose
levels are high, liver glucokinase is significantly active, which causes the
liver preferentially to trap and to store circulating glucose. When blood
glucose falls to very low levels, tissues such as liver and kidney, which
contain glucokinases but are not highly dependent on glucose, do not continue
to use the meager glucose supplies that remain available. At the same time,
tissues such as the brain, which are critically dependent on glucose, continue
to scavenge blood glucose using their low Km hexokinases, and as a
consequence their viability is protected. Under various conditions of glucose
deficiency, such as long periods between meals, the liver is stimulated to
supply the blood with glucose through the pathway of gluconeogenesis. The levels of glucose
produced during gluconeogenesis are insufficient to activate glucokinase,
allowing the glucose to pass out of hepatocytes and into the blood.
The
regulation of hexokinase and glucokinase activities is also different.
Hexokinases I, II, and III are allosterically inhibited by product (G6P)
accumulation, whereas glucokinases are not. The latter further insures liver
accumulation of glucose stores during times of glucose excess, while favoring
peripheral glucose utilization when glucose is required to supply energy to
peripheral tissues.
Phosphohexose
Isomerase:
The second
reaction of glycolysis is an isomerization, in which G6P is converted to
fructose 6-phosphate (F6P). The enzyme catalyzing this reaction is
phosphohexose isomerase (also known as phosphoglucose isomerase). The reaction
is freely reversible at normal cellular concentrations of the two hexose
phosphates and thus catalyzes this interconversion during glycolytic carbon
flow and during gluconeogenesis.
6-Phosphofructo-1-Kinase
(Phosphofructokinase-1, PFK-1):
The next
reaction of glycolysis involves the utilization of a second ATP to convert F6P
to fructose 1,6-bisphosphate (F1,6BP). This reaction is catalyzed by
6-phosphofructo-1-kinase, better known as phosphofructokinase-1 or PFK-1.
This reaction is not readily reversible because of its large positive free
energy (G0' = +5.4 kcal/mol) in the reverse direction. Nevertheless,
fructose units readily flow in the reverse (gluconeogenic) direction because of
the ubiquitous presence of the hydrolytic enzyme, fructose-1,6-bisphosphatase
(F-1,6-BPase).
The presence
of these two enzymes in the same cell compartment provides an example of a
metabolic futile cycle, which if unregulated would rapidly deplete cell energy
stores. However, the activity of these two enzymes is so highly regulated that
PFK-1 is considered to be the rate-limiting
enzyme of glycolysis and F-1,6-BPase is considered to be the rate-limiting enzyme in
gluconeogenesis.
Aldolase:
Aldolase
catalyses the hydrolysis of F1,6BP into two 3-carbon products: dihydroxyacetone
phosphate (DHAP) and glyceraldehyde 3-phosphate (G3P). The aldolase reaction
proceeds readily in the reverse direction, being utilized for both glycolysis
and gluconeogenesis.
Triose
Phosphate Isomerase: \
The two
products of the aldolase reaction equilibrate readily in a reaction catalyzed
by triose phosphate isomerase. Succeeding reactions of glycolysis utilize G3P
as a substrate; thus, the aldolase reaction is pulled in the glycolytic
direction by mass action principals.
Glyceraldehyde-3-Phosphate
Dehydrogenase:
The second
phase of glucose catabolism features the energy-yielding glycolytic reactions
that produce ATP and NADH. In the first of these reactions, glyceraldehyde-3-P
dehydrogenase (G3PDH) catalyzes the NAD+-dependent oxidation of G3P
to 1,3-bisphosphoglycerate (1,3BPG) and NADH. The G3PDH reaction is reversible,
and the same enzyme catalyzes the reverse reaction during gluconeogenesis.
Phosphoglycerate
Kinase:
The
high-energy phosphate of 1,3-BPG is used to form ATP and 3-phosphoglycerate
(3PG) by the enzyme phosphoglycerate kinase. Note that this is the only
reaction of glycolysis or gluconeogenesis that involves ATP and yet is
reversible under normal cell conditions. Associated with the phosphoglycerate
kinase pathway is an important reaction of erythrocytes, the formation of
2,3-bisphosphoglycerate, 2,3BPG (see Figure below) by the enzyme
bisphosphoglycerate mutase. 2,3BPG is an important regulator of hemoglobin's
affinity for oxygen. Note that 2,3-bisphosphoglycerate phosphatase degrades
2,3BPG to 3-phosphoglycerate, a normal intermediate of glycolysis. The 2,3BPG
shunt thus operates with the expenditure of 1 equivalent of ATP per triose
passed through the shunt. The process is not reversible under physiological
conditions.
The pathway
for 2,3-bisphosphoglycerate (2,3-BPG)
synthesis within erythrocytes. Synthesis of 2,3-BPG represents a major reaction
pathway for the consumption of glucose in erythrocytes. The synthesis of
2,3-BPG in erythrocytes is critical for controlling hemoglobin affinity for oxygen.
Note that when glucose is oxidized by this pathway the erythrocyte loses the
ability to gain 2 moles of ATP from glycolytic oxidation of 1,3-BPG to
3-phosphoglycerate via the phosphoglycerate kinase reaction.
Phosphoglycerate
Mutase and Enolase:
The remaining
reactions of glycolysis are aimed at converting the relatively low energy
phosphoacyl-ester of 3PG to a high-energy form and harvesting the phosphate as
ATP. The 3PG is first converted to 2PG by phosphoglycerate mutase and the 2PG
conversion to phosphoenoylpyruvate (PEP) is catalyzed by enolase
Pyruvate
Kinase:
The final
reaction of aerobic glycolysis is catalyzed by the highly regulated enzyme
pyruvate kinase (PK). In this strongly exergonic reaction, the high-energy
phosphate of PEP is conserved as ATP. The loss of phosphate by PEP leads to the
production of pyruvate in an unstable enol form, which spontaneously
tautomerizes to the more stable, keto form of pyruvate. This reaction
contributes a large proportion of the free energy of hydrolysis of PEP.
Anaerobic Glycolysis
http://www.youtube.com/watch?v=uCmNQQWlrc0&feature=related
Under aerobic conditions, pyruvate in most cells is
further metabolized via the TCA cycle. Under anaerobic conditions and
in erythrocytes under aerobic conditions, pyruvate is converted to lactate by
the enzyme lactate dehydrogenase (LDH), and the lactate is transported out of
the cell into the circulation. The conversion of pyruvate to lactate, under
anaerobic conditions, provides the cell with a mechanism for the oxidation of
NADH (produced during the G3PDH reaction) to NAD+; which occurs
during the LDH catalyzed reaction. This reduction is required since NAD+
is a necessary substrate for G3PDH, without which glycolysis will cease.
Normally, during aerobic glycolysis the electrons of cytoplasmic NADH are
transferred to mitochondrial carriers of the oxidative phosphorylation pathway
generating a continuous pool of cytoplasmic NAD+.
Aerobic
glycolysis generates substantially more ATP per mole of glucose oxidized than
does anaerobic glycolysis. The utility of anaerobic glycolysis, to a muscle
cell when it needs large amounts of energy, stems from the fact that the rate
of ATP production from glycolysis is approximately 100X faster than from
oxidative phosphorylation. During exertion muscle cells do not need to energize
anabolic reaction pathways. The requirement is to generate the maximum amount
of ATP, for muscle contraction, in the shortest time frame. This is why muscle
cells derive almost all of the ATP consumed during exertion from anaerobic
glycolysis.
The reactions
catalyzed by hexokinase, PFK-1 and PK all proceed with a relatively large free
energy decrease. These nonequilibrium reactions of glycolysis would be ideal
candidates for regulation of the flux through glycolysis. Indeed, in vitro
studies have shown all three enzymes to be allosterically controlled.
Regulation of
hexokinase, however, is not the major control point in glycolysis. This is due
to the fact that large amounts of G6P are derived from the breakdown of
glycogen (the predominant mechanism of carbohydrate entry into glycolysis in
skeletal muscle) and, therefore, the hexokinase reaction is not necessary.
Regulation of PK is important for reversing glycolysis when ATP is high in
order to activate gluconeogenesis. As such this enzyme catalyzed reaction is
not a major control point in glycolysis. The rate limiting step in glycolysis
is the reaction catalyzed by PFK-1.
PFK-1 is a
tetrameric enzyme that exist in two conformational states termed R and T that
are in equilibrium. ATP is both a substrate and an allosteric inhibitor of
PFK-1. Each subunit has two ATP binding sites, a substrate site and an
inhibitor site. The substrate site binds ATP equally well when the tetramer is
in either conformation. The inhibitor site binds ATP essentially only when the
enzyme is in the T state. F6P is the other substrate for PFK-1 and it also
binds preferentially to the R state enzyme. At high concentrations of ATP, the
inhibitor site becomes occupied and shifting the equilibrium of PFK-1
comformation to that of the T state decreasing PFK-1's ability to bind F6P. The
inhibition of PFK-1 by ATP is overcome by AMP which binds to the R state of the
enzyme and, therefore, stabilizes the conformation of the enzyme capable of
binding F6P. The most important allosteric regulator of both glycolysis and
gluconeogenesis is fructose
2,6-bisphosphate, F2,6BP, which is not an intermediate in glycolysis or
in gluconeogenesis.
Regulation of
glycolysis and gluconeogenesis by fructose
2,6-bisphosphate (F2,6BP). The major sites for regulation of glycolysis
and gluconeogenesis are the phosphofructokinase-1 (PFK-1) and
fructose-1,6-bisphosphatase (F-1,6-BPase) catalyzed reactions. PFK-2 is the
kinase activity and F-2,6-BPase is the phosphatase activity of the
bi-functional regulatory enzyme,
phosphofructokinase-2/fructose-2,6-bisphosphatase. PKA is cAMP-dependent
protein kinase which phosphorylates PFK-2/F-2,6-BPase turning on the
phosphatase activity. (+ve) and (-ve) refer to positive and negative
activities, respectively.
The synthesis
of F2,6BP is catalyzed by the bifunctional enzyme
phosphofructokinase-2/fructose-2,6-bisphosphatase (PFK-2/F-2,6-BPase). In the
nonphosphorylated form the enzyme is known as PFK-2 and serves to catalyze the
synthesis of F2,6BP by phosphorylating fructose 6-phosphate. The result is that
the activity of PFK-1 is greatly stimulated and the activity of F-1,6-BPase is
greatly inhibited.
Under
conditions where PFK-2 is active, fructose flow through the PFK-1/F-1,6-BPase
reactions takes place in the glycolytic direction, with a net production of
F1,6BP. When the bifunctional enzyme is phosphorylated it no longer exhibits
kinase activity, but a new active site hydrolyzes F2,6BP to F6P and inorganic
phosphate. The metabolic result of the phosphorylation of the bifunctional
enzyme is that allosteric stimulation of PFK-1 ceases, allosteric inhibition of
F-1,6-BPase is eliminated, and net flow of fructose through these two enzymes
is gluconeogenic, producing F6P and eventually glucose.
The
interconversion of the bifunctional enzyme is catalyzed by cAMP-dependent
protein kinase (PKA), which in turn is regulated by circulating peptide
hormones. When blood glucose levels drop, pancreatic insulin production falls,
glucagon secretion is stimulated, and circulating glucagon is highly increased.
Hormones such as glucagon bind to plasma membrane receptors on liver cells,
activating membrane-localized adenylate cyclase leading to an increase in the
conversion of ATP to cAMP (see diagram below). cAMP binds to the regulatory
subunits of PKA, leading to release and activation of the catalytic subunits.
PKA phosphorylates numerous enzymes, including the bifunctional
PFK-2/F-2,6-BPase. Under these conditions the liver stops consuming glucose and
becomes metabolically gluconeogenic, producing glucose to reestablish
normoglycemia.
Representative
pathway for the activation of cAMP-dependent
protein kinase (PKA). In this example glucagon binds to its'
cell-surface receptor, thereby activating the receptor. Activation of the
receptor is coupled to the activation of a receptor-coupled G-protein
(GTP-binding and hydrolyzing protein) composed of 3 subunits. Upon activation
the alpha subunit dissociates and binds to and activates adenylate cyclase.
Adenylate cylcase then converts ATP to cyclic-AMP (cAMP). The cAMP thus
produced then binds to the regulatory subunits of PKA leading to dissociation
of the associated catalytic subunits. The catalytic subunits are inactive until
dissociated from the regulatory subunits. Once released the catalytic subunits
of PKA phosphorylate numerous substrate using ATP as the phosphate donor.
Regulation of
glycolysis also occurs at the step catalyzed by pyruvate kinase, (PK). The
liver enzyme has been most studied in vitro. This enzyme is inhibited by
ATP and acetyl-CoA and is activated by F1,6BP. The inhibition of PK by ATP is
similar to the effect of ATP on PFK-1. The binding of ATP to the inhibitor site
reduces its affinity for PEP. The liver enzyme is also controlled at the level
of synthesis. Increased carbohydrate ingestion induces the synthesis of PK
resulting in elevated cellular levels of the enzyme.
A number of
PK isozymes have been described. The liver isozyme (L-type), characteristic of
a gluconeogenic tissue, is regulated via phosphorylation by PKA, whereas the
M-type isozyme found in brain, muscle, and other glucose requiring tissue is
unaffected by PKA. As a consequence of these differences, blood glucose levels
and associated hormones can regulate the balance of liver gluconeogenesis and
glycolysis while muscle metabolism remains unaffected.
In
erythrocytes, the fetal PK isozyme has much greater activity than the adult
isozyme; as a result, fetal erythrocytes have comparatively low concentrations
of glycolytic intermediates. Because of the low steady-state concentration of
fetal 1,3BPG, the 2,3BPG shunt (see diagram above) is greatly reduced in fetal
cells and little 2,3BPG is formed. Since 2,3BPG is a negative effector of
hemoglobin affinity for oxygen, fetal erythrocytes have a higher oxygen
affinity than maternal erythrocytes. Therefore, transfer of oxygen from
maternal hemoglobin to fetal hemoglobin is favored, assuring the fetal oxygen
supply. In the newborn, an erythrocyte isozyme of the M-type with comparatively
low PK activity displaces the fetal type, resulting in an accumulation of
glycolytic intermediates. The increased 1,3BPG levels activate the 2,3BPG
shunt, producing 2,3BPG needed to regulate oxygen binding to hemoglobin.
Genetic
diseases of adult erythrocyte PK are known in which the kinase is virtually
inactive. The erythrocytes of affected individuals have a greatly reduced
capacity to make ATP and thus do not have sufficient ATP to perform activities
such as ion pumping and maintaining osmotic balance. These erythrocytes have a
short half-life, lyse readily, and are responsible for some cases of hereditary hemolytic anemia.
The liver PK
isozyme is regulated by phosphorylation, allosteric effectors, and modulation
of gene expression. The major allosteric effectors are F1,6BP, which stimulates
PK activity by decreasing its Km(app) for PEP, and for the negative
effector, ATP. Expression of the liver PK gene is strongly influenced by the
quantity of carbohydrate in the diet, with high-carbohydrate diets inducing up
to 10-fold increases in PK concentration as compared to low carbohydrate diets.
Liver PK is phosphorylated and inhibited by PKA, and thus it is under hormonal
control similar to that described earlier for PFK-2.
Muscle PK (M-type)
is not regulated by the same mechanisms as the liver enzyme. Extracellular
conditions that lead to the phosphorylation and inhibition of liver PK, such as
low blood glucose and high levels of circulating glucagon, do not inhibit the
muscle enzyme. The result of this differential regulation is that hormones such
as glucagon and epinephrine favor liver gluconeogenesis by inhibiting liver
glycolysis, while at the same time, muscle glycolysis can proceed in accord
with needs directed by intracellular conditions.
Metabolic Fates of Pyruvate
Pyruvate is
the branch point molecule of glycolysis. The ultimate fate of pyruvate depends
on the oxidation state of the cell. In the reaction catalyzed by G3PDH a
molecule of NAD+ is reduced to NADH. In order to maintain the re-dox
state of the cell, this NADH must be re-oxidized to NAD+. During
aerobic glycolysis this occurs in the mitochondrial electron transport chain
generating ATP. Thus, during aerobic glycolysis ATP is generated from oxidation
of glucose directly at the PGK and PK reactions as well as indirectly by
re-oxidation of NADH in the oxidative phosphorylation pathway.
Additional NADH molecules are generated during the complete aerobic oxidation
of pyruvate in the TCA cycle. Pyruvate enters the TCA cycle
in the form of acetyl-CoA which
is the product of the pyruvate dehydrogenase reaction. The fate of pyruvate
during anaerobic glycolysis is reduction to lactate.
Lactate Metabolism
During
anaerobic glycolysis, that period of time when glycolysis is proceeding at a
high rate (or in anaerobic organisms), the oxidation of NADH occurs through the
reduction of an organic substrate. Erythrocytes and skeletal muscle (under
conditions of exertion) derive all of their ATP needs through anaerobic
glycolysis. The large quantity of NADH produced is oxidized by reducing
pyruvate to lactate. This reaction is carried out by lactate dehydrogenase,
(LDH). The lactate produced during anaerobic glycolysis diffuses from the
tissues and is transproted to highly aerobic tissues such as cardiac muscle and
liver. The lactate is then oxidized to pyruvate in these cells by LDH and the
pyruvate is further oxidized in the TCA cycle. If the energy level in these
cells is high the carbons of pyruvate will be diverted back to glucose via the
gluconeogenesis pathway.
Mammalian
cells contain two distinct types of LDH subunits, termed M and H. Combinations
of these different subunits generates LDH isozymes with different characteristics.
The H type subunit predominates in aerobic tissues such as heart muscle (as the
H4 tetramer) while the M subunit predominates in anaerobic tissues such as
skeletal muscle as the M4 tetramer). H4 LDH has a low Km for
pyruvate and also is inhibited by high levels of pyruvate. The M4 LDH enzyme
has a high Km for pyruvate and is not inhibited by pyruvate. This
suggsts that the H-type LDH is utilized for oxidizing lactate to pyruvate and
the M-type the reverse.
The content
of glucose in blood:
-
source of
blood glucose;
Blood sugar concentration, or glucose level, refers to the
amount of glucose
present in a mammal's blood.
Normally, the blood glucose level is maintained at a reference
range between about 4 and 6 mM (mmol/l). It is tightly regulated
in the human body. The normal
blood glucose level is about 90mg/100ml, which works out to 5mM (mmol/l), since
the molecular weight of glucose, C6H12O6, is
about 180 g/mol daltons.
The total amount of glucose in circulating blood is therefore about 3.3 to 7g
(assuming an ordinary adult blood volume of 5 litres, plausible for an average
adult male). Glucose levels rise after meals for an hour or two by a few grams
and are usually lowest in the morning, before the first meal of the day.
Transported via the bloodstream from the intestines or liver
to body cells, Glucose is the primary source of energy for the body's cells.
Failure to maintain blood glucose in
the normal range leads to conditions of persistently high (hyperglycemia)
or low (hypoglycemia)
blood sugar. Diabetes
mellitus, characterized by persistent hyperglycemia from any of several
causes, is the most prominent disease related to failure of blood sugar
regulation
Causes and
consequences of hypo- and hyperglycemia
Hyperglycemia, hyperglycaemia, or high blood sugar
is a condition in which an excessive amount of glucose
circulates in the blood
plasma. This is generally a blood glucose level of 10+ mmol/L
(180 mg/dl), but symptoms may not start to become noticeable until later
numbers like 15-20+ mmol/L
(270-360 mg/dl). However, chronic levels exceeding 125 mg/dl can produce organ
damage.
The origin of the term is Greek:
hyper-, meaning excessive; -glyc-, meaning sweet; and -emia,
meaning "of the blood".
Hypoglycaemia or hypoglycemia is the medical term for a pathologic
state produced by a lower than normal level of blood
glucose. The term hypoglycemia literally means "under-sweet
blood" (Gr.
hypo-, glykys, haima). The term also refers to a putative
condition that is scientifically disputed and which is perhaps more properly
considered as a part of "alternative" medicine.[neutrality
disputed] This is covered at the end of this article.
Hypoglycemia can produce a variety of
symptoms
and effects but the principal problems arise from an inadequate supply of
glucose as fuel to the brain,
resulting in impairment of function (neuroglycopenia).
Derangements of function can range from vaguely "feeling bad" to coma,
anymous seizures,
and (rarely) permanent brain damage or death. Hypoglycemia can arise from many
causes and can occur at any age. It also sometimes occurs at random.
The most common forms of moderate and severe hypoglycemia occur as a
complication of treatment of diabetes
mellitus treated with insulin
or less frequently with certain oral
medications. Hypoglycemia is usually treated by the ingestion or
administration of dextrose,
or foods quickly digestible to glucose.
Endocrinologists
(specialists in hormones, including those which regulate glucose
metabolism) typically consider the following criteria (referred to as Whipple's
triad) as proving that individual's symptoms
can be attributed to hypoglycemia:
1.
Symptoms
known to be caused by hypoglycemia
2.
Low glucose
at the time the symptoms occur
3.
Reversal or
improvement of symptoms or problems when the glucose is restored to normal
However, not everyone has accepted these suggested diagnostic criteria,
and even the level of glucose low enough to define hypoglycemia has been a
source of controversy in several contexts. For many purposes, plasma
glucose levels below 70 mg/dl or 3.9 mmol/L
are considered hypoglycemic; these issues are detailed below.
Biological and energetic value of
glycogenolysis
Glycogenolysis:
In glycogenolysis, glycogen stored in
the liver and muscles, is converted first to glucose-1- phosphate and then into
glucose-6-phosphate. Two hormones which control glycogenolysis are a peptide,
glucagon from the pancreas and epinephrine from the adrenal glands.
Glucagon is
released from the pancreas in response to low blood glucose and epinephrine is
released in response to a threat or stress. Both hormones act upon enzymes to
stimulate glycogen phosphorylase to begin glycogenolysis and inhibit glycogen
synthetase (to stop glycogenesis).
Glycogen is a
highly branched polymeric structure containing glucose as the basic monomer.
First individual glucose molecules are hydrolyzed from the chain, followed by
the addition of a phosphate group at C-1. In the next step the phosphate is
moved to the C-6 position to give glucose 6-phosphate, a cross road compound.
Glucose-6-phosphate
is the first step of the glycolysis pathway if glycogen is the carbohydrate
source and further energy is needed. If energy is not immediately needed, the
glucose-6-phosphate is converted to glucose for distribution in the blood to
various cells such as brain cells.
Glycogenolysis:
ІІ. Alternative ways of monosaccharides
metabolism.
Entry of
other carbohydrates (fructose, galactose) into the glycolytic sequence
Entry of
galactose into the glycolytic-gluconeogenetic pathway.
Galactose comes from
the splitting of lactose (galactose + glucose).
Lactose is broken
down by lactase, the sugars then freely enter the intestinal cells (not insulin
dependent).
Galactose (like all monosaccharides) must be phosphorylated in order to enter
the pathways.
In order for
galactose to enter the glycolytic pathway is must first be converted to
galactose-1-phosphate and then activated by adding UDP to make galactose-UDP.
Galactose intolerance.
Lack of the enzyme
galactokinase.
The accumulation
of galactitol will damage the eyes (cataract)
Hepatic metabolism
of fructose.
The diatery source
of fructose is the disaccharide sucrose (glucose+fructose).
FK = Fructokinase
GK/HK = Glucokinase / Hexokinase ( either one catalyzes this reaction, although
they both have a much higher affinity for Glucose, they are able to process
Fructose)
A = Aldolase A
B = Aldolase B
The minor pathway
is that of Fructose => Fructose-6-Phosphate (F-6-P)
The major pathway is that of Fructose => Fructose-1-Phosphate (F-1-P)
The
regulatory/toxic effects of fructose-1-phosphate.
Fructokinase acts
rapidly, forming Fructose-1-phosphate, but Aldolase B doesn't work as fast.
This increases the amount of Fructose-1-phosphate leading to an accumulation,
especially in the liver (since the liver is the major organ for fructose
metabolism).
Phosphate is bound to fructose, so the level of ATP decreases. This is called phosphate trapping .
Alcoholic
fermentation. Common reactions for
fermentation and glycolysis.
Difference of
these processes.
When the oxygen supply runs short in heavy or
prolonged exercise, muscles obtain most of their energy from an anaerobic
(without oxygen) process called glycolysis. Yeast cells obtain energy
under anaerobic conditions using a very similar process called alcoholic
fermentation. Glycolysis is the chemical breakdown of glucose to lactic
acid. This process makes energy available for cell activity in the form of a
high-energy phosphate compound known as adenosine triphosphate (ATP). Alcoholic
fermentation is identical to glycolysis except for the final step (Fig. 1).
In alcoholic fermentation, pyruvic acid is broken down into ethanol
and carbon dioxide. Lactic acid from glycolysis produces a feeling of tiredness;
the products of alcoholic fermentation have been used in baking and brewing for
centuries.
Both alcoholic
fermentation and glycolysis are anaerobic fermentation processes that begin
with the sugar glucose. Glycolysis requires 11 enzymes which degrade glucose to
lactic acid (Fig. 2). Alcoholic fermentation follows the same enzymatic pathway
for the first 10 steps. The last enzyme of glycolysis, lactate dehydrogenase,
is replaced by two enzymes in alcoholic fermentation. These two enzymes,
pyruvate decarboxylase and alcoholic dehydrogenase, convert pyruvic acid into
carbon dioxide and ethanol in alcoholic fermentation.
The most commonly
accepted evolutionary scenario states that organisms first arose in an
atmosphere lacking oxygen. Anaerobic fermentation is supposed to have evolved
first and is considered the most ancient pathway for obtaining energy. There
are several scientific difficulties, however, with considering fermentations as
primitive energy harvesting mechanisms produced by time and chance.
First of all, it
takes ATP energy to start the process that will only later generate a net gain
in ATP. Two ATPs are put into the glycolytic pathway for priming the reactions,
the expenditure of energy by conversion of ATP to ADP being required in the
first and third steps of the pathway (Fig. 2). A total of four ATPs are
obtained only later in the sequence, making a net gain of two ATPs for each
molecule of glucose degraded. The net gain of two ATPs is not realized until
the tenth enzyme in the series catalyzes phosphoenolpyruvate to ATP and pyruvic
acid (pyruvate). This means that neither glycolysis nor alcoholic fermentation
realizes any gain in energy (ATP) until the tenth enzymatic breakdown.
It is purely
wishful thinking to suppose that a series of 10 simultaneous, beneficial,
additive mutations could produce 10 complex enzymes to work on 10 highly
specific substances and that these reactions would occur in sequence. Enzymes
are proteins consisting of amino acids united in polypeptide chains. Their
complexity may be illustrated by the enzyme glyceraldehyde phosphate
dehydrogenase, which is the enzyme that catalyzes the oxidation of
phosphoglyceraldehyde in glycolysis and alcoholic fermentation. Glyceraldehyde
phosphate dehydrogenase consists of four identical chains, each having 330
amino acid residues. The number of different possible arrangements for the
amino acid residues of this enzyme is astronomical.
To illustrate, let
us consider a simple protein containing only 100 aim acids. There are 20
different kinds of L-amino acids in proteins, and each can be used repeatedly
in chains of 100. Therefore, they could be arranged in 20100 or 10130
different ways. Even if a hundred million billion of these (1017)
combinations could function for a given purpose, there is only one chance in 10113
of getting one of these required amino acid sequences in a small protein
consisting of 100 amino acids.
Fig. 2.
Notice that ATP is formed at two different locations above (steps 7 & 10).
Because there are 2 molecules of the substrates, there will be 2 molecules of
ATP formed at both locations, making a total of 4 molecules of ATP. Two
molecules of ATP were necessary for priming the original breakdown of glucose
(step 1). Therefore, a net of 2 molecules of ATP are recognized from the entire
breakdown of glucose pyruvate. (4 ATP formed - 2 ATP primers = 2 ATP net
overall gain.) Notice also that this MW net gain In ATP is not recognized until
phosphoenolpyruvate is broken down by pyruvate kinase to form 2 molecules of
pyruvate. This means that 10 enzymatic reactions must proceed in sequence,
before energy in the form of ATP is obtained.
There are still
other problems with the theory of evolution for alcoholic fermentation and
glycolytic pathways. It is necessary to account for the numerous complex
regulatory mechanisms which control these chemical pathways. For example,
phosphofructokinase is a regulatory enzyme which limits the rate of glycolysis.
Glycogen phosphorylase is also a regulatory enzyme; it converts glycogen to
glucose-1-phosphate and thus makes glycogen available for glycolytic breakdown.
In complex organisms there are several hormones such as somatotropin, insulin,
glucagon, glucocorticoids, adrenaline thyroxin and a host of others which
control utilization of glucose. No evolutionary mechanism has ever been
proposed to account for these control mechanisms.
In addition to the
regulators, complex cofactors are absolutely essential for glycolysis. One of
the two key ATP energy harvesting steps in glycolysis requires a dehydrogenase
enzyme acting in concert with the "hydrogen shuttle" redox reactant,
nicotinamide adenine dinucleotide (NAD+). To keep the reaction sequence going,
the reduced cofactor (NADH + H +) must be continuously regenerated
by steps later in the sequence (Fig. 2), and that requires one enzyme in
glycolysis (lactic dehydrogenase) and another (alcohol dehydrogenase) in
alcoholic fermentation. In the absence of continuously cycled NAD+,
"simple" anaerobic ATP energy harvest would be impossible.
And there are
further difficulties yet for evolutionary theory to surmount. At one point, an
intermediate in the glycolytic pathway is "stuck" with a phosphate
group (needed to make ATP) in the low energy third carbon position. A
remarkable enzyme, a "mutase" (Step 8), shifts the phosphate group to
the second carbon position—but only in the presence of pre-existent primer
amounts of an extraordinary molecule, 2,3-diphosphoglyceric acid. Actually, the
shift of the phosphate from the third to the second position using the
"mutase" and these "primer" molecules accomplishes nothing
notable directly, but it "sets up" the ATP energy-harvesting reaction
which occurs two steps later!
In summary, the
following items make an evolutionary origin for glycolysis and alcoholic
fermentation totally untenable: (1) the extreme improbability of getting even
one simple enzyme by random processes; (2) the fact that the overall net gain
in energy (ATP) is not recognized until pyruvate formation suggests that the
chemical reaction must proceed through at least 10 enzymatic steps and that
these steps of necessity must be in sequence; (3) the complex regulatory
mechanisms, cofactors, and "primers" necessary for glucose utilization
cannot be explained by evolutionary speculation.
On the other hand,
the tight fit among complex and interdependent steps—especially the way some reactions
take on meaning only in terms of reactions that occur much later in the
sequence—seems to point clearly to creation with a teleological purpose, by an
Intelligence and Power far greater than man's.
The principle
of lactic acid amount measurement in blood serum, diagnostic significance.
Consequences of lactemia.
During intense exercise, muscle and blood
lactate can rise to very high levels. This accumulation above resting levels represents
the balance of production and removal. It says nothing about whether
accumulation is due to an increased rate of production or decreased rate of
removal, or both. Similarly, if lactate concentrations in the blood do not rise
above resting levels during or immediately following exercise, it also infers
nothing about lactate or lactic acid production during that activity. It may be
that lactic acid production is several times higher than at rest but that it is
matched by its removal showing no net increase.
A common
misinterpretation is that blood lactate or even lactic acid, has a direct
detrimental effect on muscle performance. However, most researchers agree that
any negative effect on performance associated with blood lactate accumulation
is due to an increase in hydrogen ions. When lactic acid dissociates it forms
lactate and hydrogen ions - which leads to an increase in acidity. So it is not
accurate to blame either lactate or lactic acid for having a direct negative
impact on muscular performance.
The increase
in hydrogen ions and subsequent acidity of the internal environment is called acidosis.
Lactic
Acidosis
So this
unfavourable acidosis is the result of an increased concentration or accumulation
of hydrogen ions. It may seem logical to conclude then, that any increase in production
of lactic acid and hence lactate is detrimental as it will increase the
production of hydrogen ions. However, accumulation is the key
term here as an increased production of hydrogen ions (due to
an increase production of lactic acid) will have no detrimental effect if
clearance is just takes it a step further…
They suggest that lactate
production (especially if accompanied by a high capacity for lactate removal)
may be more likely to delay the onset of acidosis. The reasons
for this, amongst others, are that lactate serves to consume hydrogen ions and
allows the transport of hydrogen ions from the cell. Similarly, they maintain,
there is a wealth of research evidence to show that acidosis is caused by
reactions other than lactate production. Rogers et al. do conclude
however, that increased lactate concentration, although not causative,
coincides with cellular acidosis and remains a good indirect marker for the
onset of fatigue.
As mentioned
earlier, there has been substantial research to show that an increase concentration
of hydrogen ions and a decrease in pH (increase in acidity) within muscle or
plasma, causes fatigue. Additionally, induced acidosis can impair muscle
contractility even in non-fatigued humans and several mechanisms to explain
such effects have been provided.
Yet in the last 10
years a number of high profile papers have challenged even this most basic
assumption of fatigue. A 2006 review of these by Cairns suggests that
experiments on isolated muscle show that acidosis has little detrimental effect
or may even improve muscle performance during high-intensity exercise.
In place of
acidosis it may be inorganic phosphate that is major cause of
muscle fatigue. Recall that an inorganic phosphate is produced during the
breakdown of ATP to ADP. However, there are several limitations regarding this
phosphate hypothesis. Another proposal for a major contributor to fatigue, rather
than acidosis, is the accumulation of potassium ions in muscle interstitium.
Contrary to this
new research (which is by no means definitive) is the argument that if acidosis
plays no role in fatigue then it is surprising that alkalosis (through sodium
bicarbonate consumption for example) can improve exercise performance in events
lasting 1-10 minutes. To reconcile this, Cairns (18) hypothesizes that while
acidosis has little detrimental effect or may even improve muscle performance
in isolated muscle, severe blood plasma
acidosis may impair performance by causing a reduced central nervous system
drive to muscle.
Pentose phosphate pathway of carbohydrates metabolism
The
pentose phosphate pathway is primarily an anabolic pathway that utilizes the 6
carbons of glucose to generate 5 carbon sugars and reducing equivalents.
However, this pathway does oxidize glucose and under certain conditions can
completely oxidize glucose to CO2 and water. The primary functions
of this pathway are:
1. To generate reducing equivalents, in
the form of NADPH, for reductive biosynthesis reactions within cells.
2. To provide the cell with
ribose-5-phosphate (R5P) for the synthesis of the nucleotides and nucleic
acids.
3. Although not a significant function
of the PPP, it can operate to metabolize dietary pentose sugars derived from
the digestion of nucleic acids as well as to rearrange the carbon skeletons of
dietary carbohydrates into glycolytic/gluconeogenic intermediates.
Enzymes that
function primarily in the reductive direction utilize the NADP+/NADPH
cofactor pair as co-factors as opposed to oxidative enzymes that utilize the
NAD+/NADH cofactor pair. The reactions of fatty acid biosynthesis
and steroid biosynthesis utilize large amounts of NADPH. As a consequence,
cells of the liver, adipose tissue, adrenal cortex, testis and lactating
mammary glan have high levels of the PPP enzymes. In fact 30% of the oxidation
of glucose in the liver occurs via the PPP. Additionally, erythrocytes utilize
the reactions of the PPP to generate large amounts of NADPH used in the
reduction of glutathione (see below). The conversion of ribonucleotides to
deoxyribonucleotides (through the action of ribonucleotide reductase) requires
NADPH as the electron source, therefore, any rapidly proliferating cell needs
large quantities of NADPH.
The reactions
of the PPP operate exclusively in the cytoplasm. From this perspective it is
understandable that fatty acid synthesis (as opposed to oxidation) takes place
in the cytoplasm. The pentose phosphate pathway has both an oxidative and a
non-oxidative arm. The oxidation steps, utilizing glucose-6-phosphate (G6P) as
the substrate, occur at the beginning of the pathway and are the reactions that
generate NADPH. The reactions catalyzed by glucose-6-phosphate dehydrogenase
and 6-phosphogluconate dehydrogenase generate one mole of NADPH each for every
mole of glucose-6-phosphate (G6P) that enters the PPP.
The
non-oxidative reactions of the PPP are primarily designed to generate R5P.
Equally important reactions of the PPP are to convert dietary 5 carbon sugars
into both 6 (fructose-6-phosphate) and 3 (glyceraldehyde-3-phosphate) carbon
sugars which can then be utilized by the pathways of glycolysis.
The primary
enzymes involved in the non-oxidative steps of the PPP are transaldolase and
transketolase:
Transketolase
functions to transfer 2 carbon groups from substrates of the PPP, thus rearranging
the carbon atoms that enter this pathway. Like other enzymes that transfer 2
carbon groups, transketolase requires thiamine pyrophosphate (TPP) as a
co-factor in the transfer reaction.
Transaldolase
transfers 3 carbon groups and thus is also involved in a rearrangement of the
carbon skeletons of the substrates of the PPP. The transaldolase reaction involves
Schiff base formation between the substrate and a lysine residue in the enzyme.
The net
result of the PPP, if not used solely for R5P production, is the oxidation of
G6P, a 6 carbon sugar, into a 5 carbon sugar. In turn, 3 moles of 5 carbon
sugar are converted, via the enzymes of the PPP, back into two moles of 6
carbon sugars and one mole of 3 carbon sugar. The 6 carbon sugars can be
recycled into the pathway in the form of G6P, generating more NADPH. The 3
carbon sugar generated is glyceraldehyde-3-phsphate which can be shunted to
glycolysis and oxidized to pyruvate. Alternatively, it can be utilized by the
gluconeogenic enzymes to generate more 6 carbon sugars (fructose-6-phosphate or
glucose-6-phosphate).
Metabolic
Disorders Associated with the PPP
Diabetes,
Carbohydrate-Modified Diets, and Carbohydrate Counting
Diabetes is a
condition that alters the way the body handles carbohydrates. In terms of diet
modifications, diabetics can control blood sugar levels by appropriately
managing the carbohydrates, proteins, and fats in their meals. The amount of
carbohydrates, not necessarily the source, is the primary issue. Blood glucose
levels after a meal can be related to the process of food preparation, the
amount of food eaten, fat intake, sugar absorption, and the combination of
foods in the meal or snack.
One method of
monitoring carbohydrate levels—carbohydrate counting—assigns a certain number
of carbohydrate grams or exchanges to specific foods. Calculations are used to
determine insulin need, resulting in better control of blood glucose
levels with a larger variety of foods. Overall, diabetic diets can include
moderate amounts of sugar, as long as they are carefully monitored.
Oxidative
stress within cells is controlled primarily by the action of the peptide,
glutathione, GSH. See Specialized Products of Amino Acids for the
synthesis of GSH. GSH is a tripeptide composed of γ-glutamate, cysteine and glycine. The sulfhydryl side
chains of the cysteine residues of two glutathione molecules form a disulfide
bond (GSSG) during the course of being oxidized in reactions with various
oxides and peroxides in cells. Reduction of GSSG to two moles of GSH is the
function of glutathione reductase, an enzyme that requires coupled oxidation of
NADPH.
The cysteine
thiol of GSH plays the role in reducing oxidized thiols in other proteins.
Oxidation of 2 cysteine thiols forms a disulfide bond. Although this bond plays
a very important role in protein structure and function, inappropriately
introduced disulfides can be detrimental. Glutathione can reduce disulfides
nonenzymatically. Oxidative stress also generates peroxides that in turn can be
reduced by glutathione to generate water and an alcohol, or 2 waters if the
peroxide were hydrogen peroxide.
Regeneration
of reduced glutathione is carried out by the enzyme, glutathione reductase.
This enzyme requires the co-factor NADPH when operating in the direction of
glutathione reduction which is the thermodynamically favored direction of the
reaction.
It should be
clear that any disruption in the level of NADPH may have a profound effect upon
a cells ability to deal with oxidative stress. No other cell than the
erythrocyte is exposed to greater oxidizing conditions. After all it is the
oxygen carrier of the body.






















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
![Hypoglycaemia or hypoglycemia is the medical term for a pathologic state produced by a lower than normal level of blood glucose. The term hypoglycemia literally means "under-sweet blood" (Gr. hypo-, glykys, haima). The term also refers to a putative condition that is scientifically disputed and which is perhaps more properly considered as a part of "alternative" medicine.[neutrality disputed] This is covered at the end of this article.](http://cms.xtend-life-natural-energy.com/files/health_cond/hyperglycemia.gif){kind=link}
{kind=link}

{kind=link}
{kind=link}