1. Anatomy of the endocrine
system.
2. Anatomy of the immune
system.
3. Anatomy of the spinal
cord.
Lesson # 4
Theme 1. Anatomy of the endocrine system
ENDOCRINE GLANDS do not have the ducts, their secret gets
immediately into blood. They have prettily abundant blood supplying, and their
secret has special chemical and physiological activity. Endocrine system for
origin subdivides into glands with endodermal, mesodermal or ektodermal origin.
Glands of endodermal origin subdivide into bronchiogenic
group (thyroid, parathyroid and thymus glands) and glands developed from
epithelium of intestinal tube (endocrine part of pancreas).
Glands of mesodermal origin (interrenal system) include
interstitial cells of sexual glands and cortex of adrenal glands.
Glands of ectoderm group include hypophysis (neurogenic
group) and medulla of suprarenal glands and paraganglia.
There are certain organs which are very similar to secreting glands, but
differ from them in one essential particular, viz., they do not possess any
ducts by which their secretion is discharged. These organs are known as ductless
glands. They are capable of internal secretion—that is to say, of
forming, from materials brought to them in the blood, substances which have a
certain influence upon the nutritive and other changes going on in the body.
This secretion is carried into the blood stream, either directly by the veins
or indirectly through the medium of the lymphatics.
These glands include the thyroid, the parathyroids
and the thymus; the pituitary body and the pineal body;
the chromaphil and cortical systems to which belong the suprarenals, the
paraganglia and aortic glands, the glomus caroticum and
perhaps the glomus coccygeum. The spleen is usually included in
this list and sometimes the lymph and hemolymph nodes described
with the lymphatic system. Other glands as the liver, pancreas and sexual
glands give off internal secretions, as do the gastric and intestinal mucous
membranes.

The thyroid gland and its
relations.
The Thyroid gland is situated in
anterior neck area on level of the IV-VI cervical vertebrae and consists of
right and left lobes communicated by isthmus, which continues
upward by pyramidal portion. Thyroid gland is built by parenchyma, which
subdivides into lobuli by septa. Follicles are situated in lobules, which
contain hormones of thyroid gland: thyroxine, triiodthyronin, calcitonin. They
influence on all types of metabolism.
The Thyroid Gland (Glandula Thyreiodea)
is a highly vascular organ, situated at the front and sides of the neck; it
consists of right and left lobes connected across the middle line by a narrow
portion, the isthmus. Its weight is somewhat variable, but is usually
about
The lobes (lobuli gl. thyreoideæ) are
conical in shape, the apex of each being directed upward and lateralward as far
as the junction of the middle with the lower
third of the thyroid cartilage; the base looks downward, and is on a
level with the fifth or sixth tracheal ring. Each lobe is about
The isthmus (isthmus gl. thyreoidea) connects
together the lower thirds of the lobes; it measures about
A third lobe, of conical shape, called the pyramidal
lobe, frequently arises from the upper part of the isthmus, or from
the adjacent portion of either lobe, but most commonly the left, and ascends as
far as the hyoid bone. It is occasionally quite detached, or may be divided
into two or more parts.
A fibrous or muscular band is sometimes found attached,
above, to the body of the hyoid bone, and below to the isthmus of the gland, or
its pyramidal lobe. When muscular, it is termed the Levator glandulæ
thyreoideæ.
Small detached portions of thyroid tissue are sometimes
found in the vicinity of the lateral lobes or above the isthmus; they are
called accessory thyroid glands (glandulæ thyreoideæ
accessoriæ).

Scheme showing development of
branchial epithelial bodies. (Modified from Koh.) I, II, III,
IV. Branchial pouches.
Development.—The thyroid gland is
developed from a median diverticulum, which appears about the fourth week on
the summit of the tuberculum impar, but later is found in the furrow
immediately behind the tuberculum. It grows downward and backward as a tubular
duct, which bifurcates and subsequently subdivides into a series of cellular
cords, from which the isthmus and lateral lobes of the thyroid gland are
developed. The ultimo-branchial bodies from the fifth pharyngeal pouches are
enveloped by the lateral lobes of the thyroid gland; they undergo atrophy and
do not form true thyroid tissue. The connection of the diverticulum with the
pharynx is termed the thyroglossal duct; its continuity is subsequently
interrupted, and it undergoes degeneration, its upper end being represented by
the foramen cecum of the tongue, and its lower by the pyramidal lobe of the
thyroid gland.
Structure.—The thyroid gland is
invested by a thin capsule of connective tissue, which projects into its
substance and imperfectly divides it into masses of irregular form and size.
When the organ is cut into, it is of a brownish-red color, and is seen to be
made up of a number of closed vesicles, containing a yellow glairy fluid, and
separated from each other by intermediate connective tissue.

Section of thyroid gland of
sheep.
The vesicles of the thyroid of the adult animal are
generally closed spherical sacs; but in some young animals, e. g., young
dogs, the vesicles are more or less tubular and branched. This appearance is
supposed to be due to the mode of growth of the gland, and merely indicates
that an increase in the number of vesicles is taking place. Each vesicle is
lined by a single layer of cubical epithelium. There does not appear to be a
basement membrane, so that the epithelial cells are in direct contact with the
connective-tissue reticulum which supports the acini. The vesicles are of various
sizes and shapes, and contain as a normal product a viscid, homogeneous,
semifluid, slightly yellowish, colloid material; red corpuscles are found in it
in various stages of disintegration and decolorization, the yellow tinge being
probably due to the hemoglobin, which is thus set free from the colored
corpuscles. The colloid material contains an iodine compound, iodothyrin,
and is readily stained by eosin. According to Bensley 180 the thyroid gland prepares and secretes into the vascular channels a
substance, formed under normal conditions in the outer pole of the cell and
excreted from it directly without passing by the indirect route through the follicular
cavity. In addition to this direct mode of secretion there is an indirect mode
which consists in the condensation of the secretion into the form of droplets,
having high content of solids, and the extension of these droplets into the
follicular cavity. These droplets are formed in the same zone of the cell as
that in which the primary or direct secretion is formed.
This internal secretion of the thyroid is supposed to
contain a specific hormone which acts as a chemical stimulus to other tissues,
increasing their metabolism.
Vessels and Nerves.—The arteries
supplying the thyroid gland are the superior and inferior thyroids and
sometimes an additional branch (thyroidea ima) from the innominate artery or
the arch of the aorta, which ascends upon the front of the trachea. The
arteries are remarkable for their large size and frequent anastomoses. The veins
form a plexus on the surface of the gland and on the front of the trachea; from
this plexus the superior, middle, and inferior thyroid veins arise; the
superior and middle end in the internal jugular, the inferior in the innominate
vein. The capillary bloodvessels form a dense plexus in the connective tissue
around the vesicles, between the epithelium of the vesicles and the endothelium
of the lymphatics, which surround a greater or smaller part of the
circumference of the vesicle. The lymphatic vessels run in the
interlobular connective tissue, not uncommonly surrounding the arteries which
they accompany, and communicate with a net-work in the capsule of the gland;
they may contain colloid material. They end in the thoracic and right lymphatic
trunks. The nerves are derived from the middle and inferior cervical
ganglia of the sympathetic.
The Parathyroid gland has
pair superior parathyroid gland and inferior parathyroid gland that situated on
back surface of thyroid gland. Accessory parathyroid glands can be present.
Parathyroid gland excretes parathyroid hormone that regulates metabolism of
phosphorus and calcium.
The parathyroid glands are small brownish-red bodies, situated as
a rule between the posterior borders of the lateral lobes of the thyroid gland
and its capsule. They differ from it in structure, being composed of masses of
cells arranged in a more or less columnar fashion with numerous intervening
capillaries. They measure on an average about

Parathyroid glands.
In man, they number four as a rule; fewer than four were
found in less than 1 per cent. of over a thousand persons, but more than four
in over 33 per cent. of 122 bodies examined by Civalleri. In addition, numerous
minute islands of parathyroid tissue may be found scattered in the connective
tissue and fat of the neck around the parathyroid glands proper, and quite
distinct from them.
Development.—The parathyroid bodies
are developed as outgrowths from the third and fourth branchial pouches
A pair of diverticula arise from the fifth branchial pouch
and form what are termed the ultimo-branchial bodies: these fuse with
the thyroid gland, but probably contribute no true thyroid tissue.
Structure.—Microscopically the
parathyroids consist of intercommunicating columns of cells supported by
connective tissue containing a rich supply of blood capillaries. Most of the
cells are clear, but some, larger in size, contain oxyphil granules. Vesicles
containing colloid have been described as occurring in the parathyroid, but the
observation has not been confirmed.
No doubt the parathyroid glands produce an internal
secretion essential to the well-being of the human economy; but it is still a
matter of dispute what symptoms of disease are produced by their removal and
suppression of their secretion. Pepere believes that they show signs of
exceptional activity during pregnancy, and that parathyroid insufficiency is a
main factor in the production of tetany in infants and adults, of eclampsia,
and of certain sorts of fits. It is probable that the tetany following
parathyroidectomy is due to the accumulation of ammonium carbonate and
Endocrine part of sexual glands (testicle
and ovary)
Interstitial (Leidig) cells are situated in parenchyma of testicle. They
excrete testosteron, which influences on development of secondary sexual
signs. Corpus luteum positioned in ovaric parenchyma produces a progesteron
(it prepares a mucous membrane of the uterus membrane to embryo fixation,
detains development of new follicles and stimulates development of mammary
glands during pregnancy). Follicular epithelium excretes estrogen, which
contributes to development of primary female sexual signs (ovary and uterus)
also development of secondary female sexual signs, as growth of mammary gland,
hair according female type cetera and assists the regulation of menses.
Endocrine part of pancreas is
represented by islets of Langerhans. They produce insulin and
glucagon, that regulate metabolism of carbohydrates, regulative a sugar
contents in organism. Attached to insufficient production of these hormonal
disease sugar diabetes arises.
The pancreas is a compound racemose gland, analogous in its
structures to the salivary glands, though softer and less compactly arranged
than those organs. Its secretion, the pancreatic juice, carried by the
pancreatic duct to the duodenum, is an important digestive fluid. In addition
the pancreas has an important internal secretion, probably elaborated by the
cells of Langerhans, which is taken up by the blood stream and is concerned
with sugar metabolism. It is long and irregularly prismatic in shape; its right
extremity, being broad, is called the head, and is connected to the main
portion of the organ, or body, by a slight constriction, the
neck; while its left extremity gradually
tapers to form the tail. It is situated transversely across the
posterior wall of the abdomen, at the back of the epigastric and left
hypochondriac regions. Its length varies from 12.5 to

Transverse section through the middle
of the first lumbar vertebra, showing the relations of the pancreas.

The duodenum and pancreas.

The pancreas and duodenum from
behind.
Relations.—The Head (caput
pancreatis) is flattened from before backward, and is lodged within the
curve of the duodenum. Its upper border is overlapped by the superior part of
the duodenum and its lower overlaps the horizontal part; its right and left
borders overlap in front, and insinuate themselves behind, the descending and
ascending parts of the duodenum respectively. The angle of junction of the
lower and left lateral borders forms a prolongation, termed the uncinate
process. In the groove between the duodenum and the right lateral and lower
borders in front are the anastomosing superior and inferior pancreaticoduodenal
arteries; the common bile duct descends behind, close to the right border, to
its termination in the descending part of the duodenum.
Anterior Surface.—The greater part of
the right half of this surface is in contact with the transverse colon, only
areolar tissue intervening. From its upper part the neck springs, its
right limit being marked by a groove for the gastroduodenal artery. The lower
part of the right half, below the transverse colon, is covered by peritoneum
continuous with the inferior layer of the transverse mesocolon, and is in
contact with the coils of the small intestine. The superior mesenteric artery
passes down in front of the left half across the uncinate process; the superior
mesenteric vein runs upward on the right side of the artery and, behind the
neck, joins with the lienal vein to form the portal vein.
Posterior Surface.—The posterior surface
is in relation with the inferior vena cava, the common bile duct, the renal
veins, the right crus of the diaphragm, and the aorta.
The Neck springs from the right upper portion of the
front of the head. It is about
The Body (corpus pancreatis) is somewhat
prismatic in shape, and has three surfaces: anterior, posterior, and inferior.
The anterior surface (facies anterior) is
somewhat concave; and is directed forward and upward: it is covered by the
postero-inferior surface of the stomach which rests upon it, the two organs
being separated by the omental bursa. Where it joins the neck there is a
well-marked prominence, the tuber omentale, which abuts against the
posterior surface of the lesser omentum.
The posterior surface (facies posterior) is
devoid of peritoneum, and is in contact with the aorta, the lienal vein, the
left kidney and its vessels, the left suprarenal gland, the origin of the
superior mesenteric artery, and the crura of the diaphragm.
The inferior surface (facies inferior) is
narrow on the right but broader on the left, and is covered by peritoneum; it
lies upon the duodenojejunal flexure and on some coils of the jejunum; its left
extremity rests on the left colic flexure.
The superior border (margo superior) is blunt
and flat to the right; narrow and sharp to the left, near the tail. It
commences on the right in the omental tuberosity, and is in relation with the
celiac artery, from which the hepatic artery courses to the right just above
the gland, while the lienal artery runs toward the left in a groove along this
border.
The anterior border (margo anterior) separates
the anterior from the inferior surface, and along this border the two layers of
the transverse mesocolon diverge from one another; one passing upward over the
anterior surface, the other backward over the inferior surface.
The inferior border (margo inferior) separates
the posterior from the inferior surface; the superior mesenteric vessels emerge
under its right extremity.
The Tail (cauda pancreatis) is narrow; it
extends to the left as far as the lower part of the gastric surface of the
spleen, lying in the phrenicolienal ligament, and it is in contact with the
left colic flexure.

The pancreatic duct.
The Pancreatic Duct (ductus pancreaticus [Wirsungi];
duct of Wirsung) extends transversely from left to right through the
substance of the pancreas (1100).
It commences by the junction of the small ducts of the lobules
situated in the tail of the pancreas, and, running from left to right through
the body, it receives the ducts of the various lobules composing the gland.
Considerably augmented in size, it reaches the neck, and turning downward,
backward, and to the right, it comes into relation with the common bile duct,
which lies to its right side; leaving the head of the gland, it passes very
obliquely through the mucous and muscular coats of the duodenum, and ends by an
orifice common to it and the common bile duct upon the summit of the duodenal
papilla, situated at the medial side of the descending portion of the duodenum,
7.5 to
Development is developed in two parts,
a dorsal and a ventral. The former arises as a diverticulum from the dorsal
aspect of the duodenum a short distance above the hepatic diverticulum, and,
growing upward and backward into the dorsal mesogastrium, forms a part of the
head and uncinate process and the whole of the body and tail of the pancreas.
The ventral part appears in the form of a diverticulum from the primitive
bile-duct and forms the remainder of the head and uncinate process of the
pancreas. The duct of the dorsal part (accessory pancreatic duct)
therefore opens independently into the duodenum, while that of the ventral part
(pancreatic duct) opens with the common bile-duct. About the sixth week
the two parts of the pancreas meet and fuse and a communication is established
between their ducts. After this has occurred the terminal part of the accessory
duct, i. e., the part between the duodenum and the point of meeting of
the two ducts, undergoes little or no enlargement, while the pancreatic duct
increases in size and forms the main duct of the gland. The opening of the
accessory duct into the duodenum is sometimes obliterated, and even when it
remains patent it is probable that the whole of the pancreatic secretion is
conveyed through the pancreatic duct.

Pancreas of a human embryo at end
of sixth week.
1103– Schematic and enlarged cross-section through the body of a human
embryo in the region of the mesogastrium. Beginning of third month. (Toldt.) (See enlarged image)
At first the pancreas is directed upward and backward
between the two layers of the dorsal mesogastrium, which give to it a complete
peritoneal investment, and its surfaces look to the right and left. With the
change in the position of the stomach the dorsal mesogastrium is drawn downward
and to the left, and the right side of the pancreas is directed backward and
the left forward. The right surface becomes applied to the posterior abdominal
wall, and the peritoneum which covered it undergoes absorption, and thus, in
the adult, the gland appears to lie behind the peritoneal cavity.
Structure—In structure, the pancreas
resembles the salivary glands. It differs from them, however, in certain
particulars, and is looser and softer in its texture. It is not enclosed in a
distinct capsule, but is surrounded by areolar tissue, which dips into its
interior, and connects together the various lobules of which it is composed.
Each lobule, like the lobules of the salivary glands, consists of one of the
ultimate ramifications of the main duct, ending in a number of cecal pouches or
alveoli, which are tubular and somewhat convoluted. The minute ducts connected
with the alveoli are narrow and lined with flattened cells. The alveoli are
almost completely filled with secreting cells, so that scarcely any lumen is
visible. In some animals spindle-shaped cells occupy the center of the alveolus
and are known as the centroacinar cells of Langerhans. These are
prolongations of the terminal ducts. The true secreting cells which line the
wall of the alveolus are very characteristic. They are columnar in shape and
present two zones: an outer one, clear and finely striated next the basement
membrane, and an inner granular one next the lumen. In hardened specimens the
outer zone stains deeply with various dyes, whereas the inner zone stains
slightly. During activity the granular zone gradually diminishes in size, and
when exhausted is only seen as a small area next to the lumen. During the
resting stages it gradually increases until it forms nearly three-fourths of
the cell. In some of the secreting cells of the pancreas is a spherical mass,
staining more easily than the rest of the cell; this is termed the paranucleus,
and is believed to be an extension from the nucleus. The connective tissue
between the alveoli presents in certain parts collections of cells, which are
termed interalveolar cell islets (islands of Langerhans). The cells of these stain lightly with hematoxylin or carmine, and are
more or less polyhedral in shape, forming a net-work in which ramify many
capillaries. There are two main types of cell in the islets, distinguished as
A-cells and B-cells according to the special staining reactions of the granules
they contain. The cell islets have been supposed to produce the internal
secretion of the pancreas which is necessary for carbohydrate metabolism, but
numerous researches have so far failed to elucidate their real function.

Section of pancreas
The walls of the pancreatic duct are thin, consisting of two
coats, an external fibrous and an internal mucous; the latter is smooth, and
furnished near its termination with a few scattered follicles.
Vessels and Nerves.—The arteries of
the pancreas are derived from the lienal, and the pancreaticoduodenal branches
of the hepatic and superior mesenteric. Its veins open into the lienal
and superior mesenteric veins. Its nerves are filaments from the lienal
plexus.
The
Adrenal gland is a pair
endocrine gland, which lies on superior extremity of right and left kidneys on
level of the Th 11 – Th 12 vertebrae. Each adrenal gland has triangle shape and
has anterior surface, posterior surface and renal surface and superior margin
and medial margin, and also has the hilus and consists of cortex and medulla
matter. Cortex produces mineralocorticoids (aldosterone), glucocorticoids and
androgens. Medulla of adrenal glands produces adrenalin and noradrenalin.

Suprarenal glands viewed from the
front.

Suprarenal glands viewed from
behind.
The
Paraganglia are small agglomerations of
chromaffin cells, placed closely near abdominal aorta (aortic paraganglia) or
in thickness of sympathetic trunk (sympathetic paraganglion). Paraganglia has a
function, analogic to function of medulla suprarenal gland.
Chromaphil or chromaffin cells, so-called because they stain yellow or brownish
with chromium salts, are associated with the ganglia of the sympathetic nervous
system.
Development.—They arise in common
with the sympathetic cells from the neural crest, and are therefore ectodermal
in origin. The chromaphil and sympathetic cells are indistinguishable from one
another at the time of their migration from the spinal ganglia to the regions
occupied in the adult. Differentiation of chromaphil cells begins in embryos
about
The paraganglia are small groups of chromaphil cells
connected with the ganglia of the sympathetic trunk and the ganglia of the
celiac, renal, suprarenal, aortic and hypogastric plexuses. They are sometimes
found in connection with the ganglia of other sympathetic plexuses. None have
been found with the sympathetic ganglia associated with the branches of the
trigeminal nerve.
The aortic glands or bodies are the largest of
these groups of chromaphil cells and measure in the newborn about
The medullary portions of the suprarenal glands and the
glomus caroticum belong to the chromaphil system.
Small accessory suprarenals (glandulæ
suprarenales accessoriæ) are often to be found in the connective
tissue around the suprarenals. The smaller of these, on section, show a uniform
surface, but in some of the larger a distinct medulla can be made out.
Structure.—On section, the
suprarenal gland is seen to consist of two portions (1185): an external or cortical and an internal or medullary.
The former constitutes the chief part of the organ, and is of a deep yellow
color; the medullary substance is soft, pulpy, and of a dark red or brown
color.
The cortical portion (substantia corticalis)
consists of a fine connective-tissue net-work, in which is imbedded the
glandular epithelium. The epithelial cells are polyhedral in shape and possess
rounded nuclei; many of the cells contain coarse granules, others lipoid
globules. Owing to differences in the arrangement of the cells, three distinct
zones can be made out: (1) the zona glomerulosa, situated beneath the
capsule, consists of cells arranged in rounded groups, with here and there
indications of an alveolar structure; the cells of this zone are very granular,
and stain deeply. (2) The zona fasciculata, continuous with the zona
glomerulosa, is composed of columns of cells arranged in a radial manner; these
cells contain finer granules and in many instances globules of lipoid material.
(3) The zona reticularis, in contact with the medulla, consists of
cylindrical masses of cells irregularly arranged; these cells often contain
pigment granules which give this zone a darker appearance than the rest of the
cortex.
The medullary portion (substantia medullaris)
is extremely vascular, and consists of large chromaphil cells arranged in a
network. The irregular polyhedral cells have a finely granular cystoplasm that
are probably concerned with the secretion of adrenalin. In the meshes of the
cellular network are large anastomosing venous sinuses (sinusoids) which are in
close relationship with the chromaphil or medullary cells. In many places the
endothelial lining of the blood sinuses is in direct contact with the medullary
cells. Some authors consider the endothelium absent in places and here the
medullary cells are directly bathed by the blood. This intimate relationship
between the chromaphil cells and the blood stream undoubtedly facilitates the
discharge of the internal secretion into the blood. There is a loose meshwork
of supporting connective tissue containing non-striped muscle fibers. This
portion of the gland is richly supplied with non-medullated nerve fibers, and
here and there sympathetic ganglia are found.

Section of a part of a suprarenal
gland.
Vessels and Nerves.—The arteries
supplying the suprarenal glands are numerous and of comparatively large size;
they are derived from the aorta, the inferior phrenic, and the renal. They
subdivide into
minute branches previous to entering the cortical part of the gland,
where they break up into capillaries which end in the venous plexus of the
medullary portion.
The suprarenal vein returns the blood from the
medullary venous plexus and receives several branches from the cortical
substance; it emerges from the hilum of the gland and on the right side opens
into the inferior vena cava, on the left into the renal vein.
The lymphatics end in the lumbar glands.
The nerves are exceedingly numerous, and are derived
from the celiac and renal plexuses, and, according to Bergmann, from the
phrenic and vagus nerves. They enter the lower and medial part of the capsule,
traverse the cortex, and end around the cells of the medulla. They have
numerous small ganglia developed upon them in the medullary portion of the
gland.
In connection with the development of the medulla from the
sympathochromaffin tissue, it is to be noted that this portion of the gland
secretes a substance, adrenalin, which has a powerful influence on those
muscular tissues which are supplied by sympathetic fibers.
Glomus Caroticum (Carotid Glands; Carotid Bodies)—The carotid bodies, two in number, are situated one on either
side of the neck, behind the common carotid artery at its point of bifurcation
into the external and internal carotid trunks. They are reddish brown in color
and oval in shape, the long diameter measuring about

Section of part of human glomus
caroticum. Highly magnified. Numerous bloodvessels are seen in section among
the gland cells.
Each is invested by a fibrous capsule and consists largely of spherical or
irregular masses of cells, the masses being more or less isolated from one
another by septa which extend inward from the deep surface of the capsule. The
cells are polyhedral in shape, and each contains a large nucleus imbedded in
finely granular protoplasm, which is stained yellow by chromic salts. Numerous
nerve fibers, derived from the sympathetic plexus on the carotid artery, are
distributed throughout the organ, and a net-work of large sinusoidal
capillaries ramifies among the cells.

Section of an irregular nodule of
the glomus coccygeum. (Sertoli.) X 85. The section shows the fibrous covering
of the nodule, the bloodvessels within it, and the epithelial cells of which it
is constituted.
Glomus Coccygeum (Coccygeal Gland or Body; Luschka’s Gland)—The glomus coccygeum is placed in front of, or immediately
below, the tip of the coccyx. It is about
It consists of irregular masses of round or polyhedral
cells, the cells of each mass being grouped around a dilated sinusoidal
capillary vessel. Each cell contains a large round or oval nucleus, the protoplasm
surrounding which is clear, and is not stained by chromic salts.
The
Hypophysis (pituitary body) is a small
reddish-gray body, about

The hypophysis cerebri in
position. Shown in sagittal section.

Median sagittal through the
hypophysis of an adult monkey. Semidiagrammatic.
The hypophysis consists of an anterior and a posterior lobe,
which differ from one another in their mode of development and in their
structure (1181). The anterior lobe is the larger and is somewhat kidney-shaped,
the concavity being directed backward and embracing the posterior lobe. It
consists of a pars anterior and a pars intermedia, separated from each other by
a narrow cleft, the remnant of the pouch or diverticulum. The pars anterior is
extremely vascular and consists of epithelial cells of varying size and shape,
arranged in cord-like trabeculæ or alveoli and separated by large, thin-walled
bloodvessels. The pars intermedia is a thin lamina closely applied to the body
and neck of the posterior lobe and extending onto the neighboring parts of the
brain; it contains few bloodvessels and consists of finely granular cells
between which are small masses of colloid material. The pars intermedia in
spite of the fact that it arises in common with the pars anterior from the
ectoderm of the primitive buccal cavity is often considered as a part of the
posterior lobe which arises from the floor of the third ventricle of the brain.
Although of nervous origin the posterior lobe contains no nerve cells or
fibers. It consists of neuroglia cells and fibers and is invaded by columns
which grow into it from the pars intermedia; imbedded in it are large quantities
of a colloid substance histologically similar to that found in the thyroid
gland. In certain of the lower vertebrates, e.g., fishes, nervous
structures are present, and the lobe is of large size.

Vertical sections of the heads of
early embryos of the rabbit. Magnified. (From Mihalkovics.) A. From an
embryo
From the pars intermedia a substance, no doubt an internal
secretion, causes constriction of the bloodvessels with rise of arterial
blood-pressure. This substance seems to have a stimulating effect on most of
the smooth muscles, acting directly upon the muscle causing contraction. It
also increases the secretion of the urine; of the mammary glands when in
functional activity; and of the cerebrospinal fluid. Extracts of this lobe also
influence the general metabolism of the carbohydrates by accelerating the
process of glycogenolysis in the liver.
The pars anterior exercises a stimulating effect on the
growth of the skeleton and probably on connective tissues in general.
Enlargement of the hypophysis and of the cavity of the sella
turcica are found in the rare disease acromegaly, which is characterized
by gradual enlargement of the face, hands, and feet, with headache and often a
peculiar type of blindness. This blindness is due to the pressure of the
enlarging hypophysis on the optic chiasma.
Development of the Hypophysis Cerebri.—This
in the adult consists of a large anterior, consisting of the pars anterior and
the pars intermedia, and a small posterior lobe: the former is derived from the
ectoderm of the stomodeum, the latter from the floor of the fore-brain. About
the fourth week there appears a pouchlike diverticulum of the ectodermal lining
of the roof of the stomodeum. This diverticulum, pouch of Rathke (1182), is the rudiment of the anterior lobe of the hypophysis; it extends
upward in front of the cephalic end of the notochord and the remnant of the
buccopharyngeal membrane, and comes into contact with the under surface of the
fore-brain. It is then constricted off to form a closed vesicle, but remains
for a time connected to the ectoderm of the stomodeum by a solid cord of cells.
Masses of epithelial cells form on either side and in the front wall of the
vesicle, and by the growth between these of a stroma from the mesoderm the
development of the anterior lobe is completed. The upwardly directed hypophyseal
involution becomes applied to the antero-lateral aspect of a downwardly
directed diverticulum from the base of the fore-brain (page 744). This
diverticulum constitutes the future infundibulum in the floor of the third
ventricle while its inferior extremity becomes modified to form the posterior
lobe of the hypophysis. In some of the lower animals the posterior lobe
contains nerve cells and nerve fibers, but in man and the higher vertebrates
these are replaced by connective tissue. A canal, craniopharyngeal canal,
is sometimes found extending from the anterior part of the fossa hypophyseos of
the sphenoid bone to the under surface of the skull, and marks the original
position of Rathke’s pouch; while at the junction of the septum of the nose
with the palate traces of the stomodeal end are occasionally present
The Pineal
body (epiphysis) is a small reddish-gray body, about
In
early life it has a glandular structure which reaches its greatest development
at about the seventh year. Later, especially after puberty, the glandular
tissue gradually disappears and is replaced by connective tissue.
Structure.—The pineal body is destitute of nervous substance, and consists of
follicles lined by epithelium and enveloped by connective tissue. These
follicles contain a variable quantity of gritty material, composed of phosphate
and carbonate of calcium, phosphate of magnesium and ammonia, and a little
animal matter.
It
contains a substance which if injected intravenously causes fall of
blood-pressure. It seems probable that the gland furnishes an internal
secretion in children that inhibits the development of the reproductive glands
since the invasion of the gland in children, by pathological growths which
practically destroy the glandular tissue, results in accelerated development of
the sexual organs, increased growth of the skeleton and precocious mentality.
Theme 2. Anatomy of the immune system
The
Thymus is a central organ of immune system,
which is situated in anterior mediastinum on level of the 4th ribs behind
manubrium sterni. Behind thymus pericardium is situated. Thymus gland consists
of lobes - right and left, which have the lobule that built by cortex and
medulla of thymus gland. In medulla Т-lymphocyte matter acquire that
peculiarities which contribute to protective function.
The thymus is a temporary organ, attaining its largest size at
the time of puberty (Hammar), when it ceases to grow, gradually dwindles, and
almost disappears. If examined when its growth is most active, it will be found
to consist of two lateral lobes placed in close contact along the middle line,
situated partly in the thorax, partly in the neck, and extending from the
fourth costal cartilage upward, as high as the lower border of the thyroid
gland. It is covered by the sternum, and by the origins of the Sternohyoidei
and Sternothyreoidei. Below, it rests upon the pericardium, being separated
from the aortic arch and great vessels by a layer of fascia. In the neck it
lies on the front and sides of the trachea, behind the Sternohyoidei and
Sternothyreoidei. The two lobes generally differ in size; they are occasionally
united, so as to form a single mass; and sometimes separated by an intermediate
lobe. The thymus is of a pinkish-gray color, soft, and lobulated on its
surfaces. It is about

The thymus of a full-time fetus, exposed in
situ.
Development.—The thymus appears in the
form of two flask-shaped entodermal diverticula, which arise, one on either
side, from the third branchial pouch (1175), and extend lateralward and backward into the surrounding mesoderm in
front of the ventral aortæ. Here they meet and become joined to one
another by connective tissue, but there is never any fusion of the thymus
tissue proper. The pharyngeal opening of each diverticulum is soon obliterated,
but the neck of the flask persists for some time as a cellular cord. By further
proliferation of the cells lining the flask, buds of cells are formed, which
become surrounded and isolated by the invading mesoderm. In the latter,
numerous lymphoid cells make their appearance, and are agregated to form
lymphoid follicles. These lymphoid cells are probably derivatives of the
entodermal cells which lined the original diverticula and their subdivisions.
Additional portions of thymus tissue are sometimes developed from the fourth
branchial pouches. Thymus continues to grow until the time of puberty and then
begins to atrophy.

Minute structure of thymus. Follicle of
injected thymus from calf, four days old, slightly diagrammatic, magnified
about 50 diameters. The large vessels are disposed in two rings, one of which
surrounds the follicle, the other lies just within the margin of the medulla.
(Watney.) A and B. From thymus of camel, examined without
addition of any reagent. Magnified about 400 diameters. A. Large
colorless cell, containing small oval masses of hemoglobin. Similar cells are
found in the lymph glands, spleen, and medulla of bone. B. Colored blood corpuscles.
Structure.—Each lateral lobe is
composed of numerous lobules held together by delicate areolar tissue; the
entire gland being enclosed in an investing capsule of a similar but denser
structure. The primary lobules vary in size from that of a pin’s head to that
of a small pea, and are made up of a number of small nodules or follicles,
which are irregular in shape and are more or less fused together, especially
toward the interior of the gland. Each follicle is from 1 to
Each follicle is surrounded by a vascular plexus, from which
vessels pass into the interior, and radiate from the periphery toward the
center, forming a second zone just within the margin of the medullary portion.
In the center of the medullary portion there are very few vessels, and they are
of minute size.
Watney has made the important observation that hemoglobin is
found in the thymus, either in cysts or in cells situated near to, or forming
part of, the concentric corpuscles. This hemo globin occurs as granules or as
circular masses exactly resembling colored blood corpuscles. He has also
discovered, in the lymph issuing from the thymus, similar cells to those found
in the gland, and, like them, containing hemoglobin in the form of either
granules or masses. From these facts he arrives at the conclusion that the
gland is one source of the colored blood corpuscles. More recently Schaffer has
observed actual nucleated red-blood corpuscles in the thymus. The function of
the thymus is obscure. It seems to furnish during the period of growth an
internal secretion concerned with some phases of body metabolism, especially that
of the sexual glands.
Vessels and Nerves.—The arteries
supplying the thymus are derived from the internal mammary, and from the
superior and inferior thyroids. The veins end in the left innominate
vein, and in the thyroid veins. The lymphatics are described on page
698. The nerves are exceedingly minute; they are derived from the vagi
and sympathetic. Branches from the descendens hypoglossi and phrenic reach the
investing capsule, but do not penetrate into the substance of the gland.
The spleen (Lien)
is situated principally in the left hypochondriac region, but its
superior extremity extends into the epigastric region; it lies between the
fundus of the stomach and the diaphragm. It is the largest of the ductless
glands, and is of an oblong, flattened form, soft, of very friable consistence,
highly vascular, and of a dark purplish color.
Development.—The spleen appears
about the fifth week as a localized thickening of the mesoderm in the dorsal
mesogastrium above the tail of the pancreas. With the change in position of the
stomach the spleen is carried to the left, and comes to lie behind the stomach
and in contact with the left kidney. The part of the dorsal mesogastrium which
intervened between the spleen and the greater curvature of the stomach forms
the gastrosplenic ligament.
Relations.—The diaphragmatic
surface (facies diaphragmatica; external or phrenic surface) is
convex, smooth, and is directed upward, backward, and to the left, except at
its upper end, where it is directed slightly medialward. It is in relation with
the under surface of the diaphragm, which separates it from the ninth, tenth,
and eleventh ribs of the left side, and the intervening lower border of the
left lung and pleura.

The visceral surface of the spleen.
The visceral surface (1188) is divided by a ridge into an anterior or gastric and a posterior
or renal portion.
The gastric surface (facies gastrica), which
is directed forward, upward, and medialward, is broad and concave, and is in
contact with the posterior wall of the stomach; and below this with the tail of
the pancreas. It presents near its medial border a long fissure, termed the
hilum. This is pierced by several irregular apertures, for the entrance and
exit of vessels and nerves.
The renal surface (facies renalis) is directed
medialward and downward. It is somewhat flattened, is considerably narrower
than the gastric surface, and is in relation with the upper part of the
anterior surface of the left kidney and occasionally with the left suprarenal
gland.
The superior extremity (extremitas superior)
is directed toward the vertebral column, where it lies on a level with the
eleventh thoracic vertebra. The lower extremity or colic surface
(extremitas inferior) is flat, triangular in shape, and rests upon the
left flexure of the colon and the phrenicocolic ligament, and is generally in
contact with the tail of the pancreas. The anterior border (margo
anterior) is free, sharp, and thin, and is often notched, especially below;
it separates the diaphragmatic from the gastric surface. The posterior
border (margo posterior), more rounded and blunter than the
anterior, separates the renal from the diaphragmatic surface; it corresponds to
the lower border of the eleventh rib and lies between the diaphragm and left
kidney. The intermediate margin is the ridge which separates the renal and
gastric surfaces. The inferior border (internal border) separates
the diaphragmatic from the colic surface.
The spleen is almost entirely surrounded by peritoneum,
which is firmly adherent to its capsule. It is held in position by two folds of
this membrane. One, the phrenicolienal ligament, is derived from the
peritoneum, where the wall of the general peritoneal cavity comes into contact
with the omental bursa between the left kidney and the spleen; the lienal
vessels pass between its two layers (1039). The other fold, the gastrolienal ligament, is also formed of two
layers, derived from the general cavity and the omental respectively, where
they meet between the spleen and stomach (1039); the short gastric and left gastroepiploic branches of the lienal artery
run between its two layers. The lower end of the spleen is supported by the
phrenicocolic ligament (see page 1155).
The size and weight of the spleen are liable to very extreme
variations at different periods of life, in different individuals, and in the
same individual under different conditions. In the adult it is usually
about
Frequently in the neighborhood of the spleen, and especially
in the gastrolienal ligament and greater omentum, small nodules of splenic
tissue may be found, either isolated or connected to the spleen by thin bands of
splenic tissue. They are known as accessory spleens (lien
accessorius; supernumerary spleen). They vary in size from that of a pea to
that of a plum.
Structure.—The spleen is invested
by two coats: an external serous and an internal fibroelastic coat.
The external or serous coat (tunica serosa)
is derived from the peritoneum; it is thin, smooth, and in the human subject
intimately adherent to the fibroelastic coat. It invests the entire organ,
except at the hilum and along the lines of reflection of the phrenicolienal and
gastrolienal ligaments.
The fibroelastic coat (tunica albuginea)
invests the organ, and at the hilum is reflected inward upon the vessels in the
form of sheaths. From these sheaths, as well as from the inner surface of the
fibroelastic coat, numerous small fibrous bands, trabeculæ (1189), are given off in all directions; these uniting, constitute the
frame-work of the spleen. The spleen therefore consists of a number of small
spaces or areolæ, formed by the trabeculæ; in these
areolæ is contained the splenic pulp.
The fibroelastic coat, the sheaths of the vessels, and the
trabeculæ, are composed of white and yellow elastic fibrous tissues, the latter
predominating. It is owing to the presence of the elastic tissue that the
spleen possesses a considerable amount of elasticity, which allows of the very
great variations in size that it presents under certain circumstances. In
addition to these constituents of this tunic, there is found in man a small
amount of non-striped muscular fiber; and in some mammalia, e. g., dog,
pig, and cat, a large amount, so that the trabeculæ appear to consist
chiefly of muscular tissue.

Transverse section of the spleen, showing the
trabecular tissue and the splenic vein and its tributaries.

Transverse section of the human spleen, showing
the distribution of the splenic artery and its branches.
The splenic pulp (pulpa lienis) is a soft mass
of a dark reddish-brown color, resembling grumous blood; it consists of a fine
reticulum of fibers, continuous with those of the trabeculæ, to which are
applied flat, branching cells. The meshes of the reticulum are filled with
blood, in which, however, the white corpuscles are found to be in larger
proportion than they are in ordinary blood. Large rounded cells, termed splenic
cells, are also seen; these are capable of ameboid movement, and often
contain pigment and red-blood corpuscles in their interior. The cells of the
reticulum each possess a round or oval nucleus, and like the splenic cells,
they may contain pigment granules in their cytoplasm; they do not stain deeply
with carmine, and in this respect differ from the cells of the Malpighian
bodies. In the young spleen, giant cells may also be found, each containing
numerous nuclei or one compound nucleus. Nucleated red-blood corpuscles have
also been found in the spleen of young animals.
Bloodvessels of the Spleen.—The lienal
artery is remarkable for its large size in proportion to the size of the
organ, and also for its tortuous course. It divides into six or more branches,
which enter the hilum of the spleen and ramify throughout its substance (1190), receiving sheaths from an involution of the external fibrous tissue.
Similar sheaths also invest the nerves and veins.
Each branch runs in the transverse axis of the organ, from
within outward, diminishing in size during its transit, and giving off in its
passage smaller branches, some of which pass to the anterior, others to the
posterior part. These ultimately leave the trabecular sheaths, and terminate in
the proper substance of the spleen in small tufts or pencils of minute
arterioles, which open into the interstices of the reticulum formed by the
branched sustentacular cells. Each of the larger branches of the artery
supplies chiefly that region of the organ in which the branch ramifies, having
no anastomosis with the majority of the other branches.
The arterioles, supported by the minute
trabeculæ, traverse the pulp in all directions in bundles (pencilli)
of straight vessels. Their trabecular sheaths gradually undergo a
transformation, become much thickened, and converted into adenoid tissue; the
bundles of connective tissue becoming looser and their fibrils more delicate,
and containing in their interstices an abundance of lymph corpuscles (

Transverse section of a portion of the spleen.
The altered coat of the arterioles, consisting of adenoid
tissue, presents here and there thickenings of a spheroidal shape, the lymphatic
nodules (Malpighian bodies of the spleen). These bodies vary in size
from about
The arterioles end by opening freely into the splenic pulp;
their walls become much attenuated, they lose their tubular character, and the
endothelial cells become altered, presenting a branched appearance, and
acquiring processes which are directly connected with the processes of the
reticular cells of the pulp (1192). In this manner the vessels end, and the blood flowing through them
finds its way into the interstices of the reticulated tissue of the splenic
pulp. Thus the blood passing through the spleen is brought into intimate
relation with the elements of the pulp, and no doubt undergoes important
changes.
After these changes have taken place the blood is collected
from the interstices of the tissue by the rootlets of the veins, which begin
much in the same way as the arteries end. The connective-tissue corpuscles of
the pulp arrange themselves in rows, in such a way as to form an elongated
space or sinus. They become elongated and spindle-shaped, and overlap each
other at their extremities, and thus form a sort of endothelial lining of the
path or sinus, which is the radicle of a vein. On the outer surfaces of these
cells are seen delicate transverse lines or markings, which are due to minute
elastic fibrillæ arranged in a circular manner around the sinus. Thus the
channel obtains an external investment, and gradually becomes converted into a
small vein, which after a short course acquires a coat of ordinary connective
tissue, lined by a layer of flattened epithelial cells which are continuous
with the supporting cells of the pulp.

Section of the spleen, showing the termination
of the small bloodvessels.
The smaller veins unite to form larger ones; these do not
accompany the arteries, but soon enter the trabecular sheaths of the capsule,
and by their junction form six or more branches, which emerge from the hilum,
and, uniting, constitute the lienal vein, the largest radicle of the portal
vein. The veins are remarkable for their numerous anastomoses, while the
arteries hardly anastomose at all.
Theme 3. Anatomy of the spinal cord.
Nervous system may be divided into two parts, central and peripheral, also somatic and autonomic. The central nervous system consists of the brain [encephalon],
contained within the cranium, and the medulla
spinalis or spinal cord, lodged
in the vertebral canal. The peripheral
nervous system consists of a series of cranial and spinal nerves. Autonomic
nervous system has a parasympathetic and sympathetic parts.
Spinal cord
is located in vertebral canal and extends inferiorly from the position of the foramen
magnum of the occipital bone to the level of the first-second lumbar vertebra.
The terminal portion of the spinal cord is called the conus medullaris and filum
terminale extends inferiorly from it to the level second coccyx vertebrae.
The cervical enlargement is
located
at the level of 6th cervical vertebrae, and the lumbosacral enlargement
at the level of 12th thoracic vertebrae. The
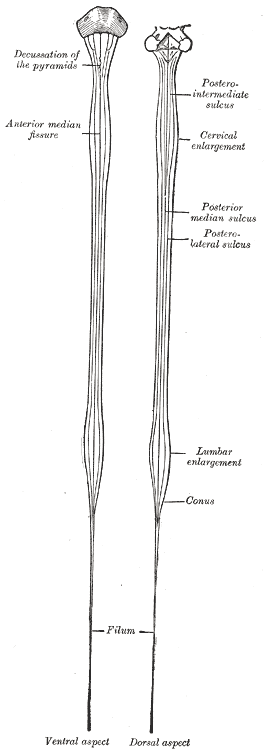
Diagrams of the medulla spinalis.
The medulla
spinalis or spinal cord forms the elongated, nearly cylindrical,
part of the central nervous system which occupies the upper two-thirds of the
vertebral canal. Its average length in the male is about
The
position of the medulla spinalis varies with the movements of the vertebral
column, its lower extremity being drawn slightly upward when the column is
flexed. It also varies at different periods of life; up to the third month of
fetal life the medulla spinalis is as long as the vertebral canal, but from
this stage onward the vertebral column elongates more rapidly than the medulla
spinalis, so that by the end of the fifth month the medulla spinalis terminates
at the base of the sacrum, and at birth about the third lumbar vertebra.

Cauda equina and filum terminale
seen from behind. The dura mater has been opened and spread out, and the
arachnoid has been removed.
The
medulla spinalis does not fill the part of the vertebral canal in which it
lies; it is ensheathed by three protective membranes, separated from each other
by two concentric spaces. The three membranes are named from without inward, the
dura mater, the arachnoid, and the pia mater. The dura
mater is a strong, fibrous membrane which forms a wide, tubular sheath;
this sheath extends below the termination of the medulla spinalis and ends in a
pointed cul-de-sac at the level of the lower border of the second sacral
vertebra. The dura mater is separated from the wall of the vertebral canal by
the epidural cavity, which contains a quantity of loose areolar tissue
and a plexus of veins; between the dura mater and the subjacent arachnoid is a
capillary interval, the subdural cavity, which contains a small quantity
of fluid, probably of the nature of lymph. The arachnoid is a thin,
transparent sheath, separated from the pia mater by a comparatively wide
interval, the subarachnoid cavity, which is filled with cerebrospinal
fluid. The pia mater closely invests the medulla spinalis and sends
delicate septa into its substance; a narrow band, the ligamentum
denticulatum, extends along each of its lateral surfaces and is attached by
a series of pointed processes to the inner surface of the dura mater.

Sagittal section of vertebral
canal to show the lower end of the medulla spinalis and the filum terminale. Li,
Lv. First and fifth lumbar vertebræ. Sii. Second sacral
vertebra. 1. Dura mater. 2. Lower part of tube of dura mater. 3. Lower
extremity of medulla spinalis. 4. Intradural, and 5, Extradural portions of
filum terminale. 6. Attachment of filum terminale to first segment of coccyx.
Thirty-one
pairs of spinal nerves spring from the medulla spinalis, each nerve having an
anterior or ventral, and a posterior or dorsal root, the latter being
distinguished by the presence of an oval swelling, the spinal ganglion,
which contains numerous nerve cells. Each root consists of several bundles of
nerve fibers, and at its attachment extends for some distance along the side of
the medulla spinalis. The pairs of spinal nerves are grouped as follows:
cervical 8, thoracic 12, lumbar 5, sacral 5, coccygeal 1, and, for convenience
of description, the medulla spinalis is divided into cervical, thoracic, lumbar
and sacral regions, corresponding with the attachments of the different groups of
nerves.
Enlargements.—The
medulla spinalis is not quite cylindrical, being slightly flattened from before
backward; it also presents two swellings or enlargements, an upper or cervical,
and a lower or lumbar.
The
cervical enlargement is the more pronounced, and corresponds with the
attachments of the large nerves which supply the upper limbs. It extends from
about the third cervical to the second thoracic vertebra, its maximum
circumference (about
The
lumbar enlargement gives attachment to the nerves which supply the lower
limbs. It commences about the level of the ninth thoracic vertebra, and reaches
its maximum circumference, of about
The
Anterior Median Fissure (fissura mediana anterior) has an average
depth of about
The Posterior Median Sulcus (sulcus medianus posterior) is
very shallow; from it a septum of neuroglia reaches rather more than half-way
into the substance of the medulla spinalis; this septum varies in depth from 4
to
On either side of the posterior median sulcus, and at a short distance
from it, the posterior nerve roots are attached along a vertical furrow named
the posterolateral sulcus. The portion of the medulla spinalis which
lies between this and the posterior median sulcus is named the posterior
funiculus. In the cervical and upper thoracic regions this funiculus
presents a longitudinal furrow, the postero-intermediate sulcus; this
marks the position of a septum which extends into the posterior funiculus and
subdivides it into two fasciculi—a medial, named the fasciculus gracilis
(tract of Goll); and a lateral, the fasciculus cuneatus (tract
of Burdach). The portion of the medulla spinalis which
lies in front of the posterolateral sulcus is termed the antero-lateral
region. The anterior nerve roots, unlike the posterior, are not attached in
linear series, and their position of exit is not marked by a sulcus. They arise
by separate bundles which spring from the anterior column of gray substance
and, passing forward through the white substance, emerge over an area of some
slight width. The most lateral of these bundles is generally taken as a
dividing line which separates the antero-lateral region into two parts, viz.,
an anterior funiculus, between the anterior median fissure and the most
lateral of the anterior nerve roots; and a lateral funiculus, between
the exit of these roots and the postero-lateral sulcus. In the upper part of
the cervical region a series of nerve roots passes outward through the lateral
funiculus of the medulla spinalis; these unite to form the spinal portion of
the accessory nerve, which runs upward and enters the cranial cavity through
the foramen magnum.
Gray Substance (substantia grisea centralis).—The
gray substance consists of two symmetrical portions, one in each half of the
medulla spinalis: these are joined across the middle line by a transverse
commissure of gray substance, through which runs a minute canal, the central
canal, just visible to the naked eye. In a transverse section each half of
the gray substance is shaped like a comma or crescent, the concavity of which
is directed laterally; and these, together with the intervening gray
commissure, present the appearance of the letter H. An imaginary coronal plane
through the central canal serves to divide each crescent into an anterior
or ventral, and a posterior or dorsal column.
The Anterior Column (columna anterior; anterior cornu), directed
forward, is broad and of a rounded or quadrangular shape. Its posterior part is
termed the base, and its anterior part the head, but these are not
differentiated from each other by any well-defined constriction. It is
separated from the surface of the medulla spinalis by a layer of white
substance which is traversed by the bundles of the anterior nerve roots. In the
thoracic region, the postero-lateral part of the anterior column projects
lateralward as a triangular field, which is named the lateral column (columna
lateralis; lateral cornu).
The Posterior Column (columna posterior; posterior cornu)
is long and slender, and is directed backward and lateralward: it reaches
almost as far as the posterolateral sulcus, from which it is separated by a thin
layer of white substance, the tract of Lissauer. It consists of a base,
directly continuous with the base of the anterior horn, and a neck or
slightly constricted portion, which is succeeded by an oval or fusiform area,
termed the head, of which the apex approaches the posterolateral
sulcus. The apex is capped by a V-shaped or crescentic mass of translucent,
gelatinous neuroglia, termed the substantia gelatinosa of Rolando, which
contains both neuroglia cells, and small nerve cells. Between the anterior and
posterior columns the gray substance extends as a series of processes into the
lateral funiculus, to form a net-work called the formatio reticularis.
The anterior median fissure (fissura mediana anterior) and right and
left anterolateral sulcuses located on anterior surface of the spinal cord. Posterior median sulcus (sulcus medianus posterior) and also
right
and
left posterolateral sulcus located
on back surface of the spinal cord.
Each
spinal nerve is attached to the medulla spinalis by two roots, an anterior or ventral (motor and sympathetic fibbers), and a posterior or dorsal, the being characterized by the
presence of a ganglion, the spinal
ganglion (with sensory pseudounipolar cells).

A spinal nerve with its anterior
and posterior roots.
After
emerging from the intervertebral foramen, each spinal nerve gives off a small meningeal branch which supplies the
dura mater, and an anterior or ventral,
posterior or dorsal divisions,
also white and gray communicating branches for nearest sympathetic ganglion (in
thoracic-lumbar part).
levels.
The
medulla spinalis is ensheathed by three protective membranes, separated from
each other by two concentric spaces:
1. The Dura
mater
2. The Arachnoid
3. The Pia mater
The
dura mater is separated from the wall of the vertebral canal
by
the epidural cavity, which contains
a quantity of fat tissue and a plexus of veins. Between the dura mater and the
subjacent arachnoid is a capillary interval, the subdural cavity, which contains a small quantity of fluid, probably
of the nature of lymph. The arachnoid
is a thin, transparent sheath, separated from the pia mater by a comparatively
wide interval, the subarachnoid cavity,
which is filled with cerebrospinal fluid. The pia mater closely invests the medulla spinalis and sends delicate
septa into its substance; a narrow band, the ligamentum denticulatum, extends along each of its lateral surfaces
and is attached by a series of pointed processes to the inner surface of the
dura mater.
|
Structure |
Location/Description |
Notes |
|
arachnoid mater |
intermediate one of the
three layers of meninges |
|
|
denticulate
ligament |
a lateral extension of pia
mater from the spinal cord |
denticulate ligament
attaches to the dura mater to anchor the spinal cord; it forms a scalloped
free border; there are 2 (one on each side) |
|
dura mater |
outermost of the meningeal layers
covering the brain and spinal cord |
"tough mother"; it
is the most durable of the meninges and provides support and protection for
the brain and spinal cord; two types are described which differ in structure:
cranial and spinal |
|
dura mater, spinal |
outermost covering of the
spinal cord, it forms the dural sac containing the spinal cord within
vertebral canal |
dural sac ends at S2,
coccygeal ligament (filum terminale externum) continues inferiorly to attach
to coccyx |
|
epidural fat |
loose connective tissue
within the epidural space |
|
|
epidural space |
the space external to the
sac of spinal dura mater within the vertebral canal |
the epidural space contains
epidural fat and the internal vertebral plexus of veins which is valveless
(clinically relevant) |
|
filum terminale
internum |
thread-like extension of the
pia mater from the conus medullaris of the spinal cord |
filum terminale internum is
best seen between vertebral levels L2 and S2; it becomes enclosed within the filum
terminale externum |
|
filum terminale
externum |
thread-like extension of the
dura mater below the end of the dural sac at S2 |
it attaches to the coccyx;
also known as the coccygeal ligament |
|
meninges |
three layers of connective
tissue covering the brain and spinal cord; dura mater, arachnoid mater, and
pia mater |
meninges provide protection
and nourishment of the brain, brainstem and spinal cord |
|
pia mater |
delicate membrane that lies
on surface of the brain and spinal cord |
"delicate mother",
it is the most delicate of the meninges; this layer faithfully follows all
surface contours of the brain and spinal cord; pia mater has 2
specializations: denticulate ligament and filum terminale internum |
|
subdural space |
the space between the dura
mater and the arachnoid mater |
this is a potential space
only; the pressure of CSF in the subarachnoid space pushes arachnoid against
dura |
|
subarachnoid space |
the space between the
arachnoid and the pia mater |
subarachnoid space contains
cerebrospinal fluid and spider web-like filaments |
|
cauda equina |
dorsal and ventral roots of
all spinal nerves inferior to L1 |
lies within the lumbar
cistern |
|
conus medullaris |
cone-shaped inferior end of
the spinal cord; located at vertebral level L1 |
at birth, the conus
medullaris is at the level of L2/L3 |
|
cervical
enlargement |
vertebral level C4 through
T1 |
created by the rootlets of
spinal nerves C5-T1 that form the brachial plexus |
|
lumbrosacral enlargement |
vertebral level T11 through
L1 |
Created by the rootlets of
spinal nerves L1-S4 that form the lumbosacral plexus |
Spinal
cord consists of the gray substance
that is surrounded by the white
substance. Posterior median sulcus passes to the gray substance and divides
by the white substance into two parts. Anterior median fissure does not pass to
the gray substance so it is white commisura that connect anterior symmetrical
regions of the white matter.
The gray substance is
largely composed of nervous cell bodies. The gray substance consists of
anterior, posterior and lateral columns (only in thoracic-lumbar portion) also
central intermediate
zone round
central canal. In transverse section columns are looking as horns, so they
differ anterior, posterior and lateral horns.

Transverse section of the medulla
spinalis in the mid-thoracic region.
Anterior horns contain
motor cells that arranged in 5 motor nuclei (nucleus anterior [medial
and lateral], central nucleus and posterior [medial and lateral] nuclei). Their
axons form anterior roots that pass
with spinal nerves to the skeletal muscles of the trunk and limbs. Posterior
horns contain intermediate cells. They receive impulse from sensory cells and
carry them to the another cell. Intermediate cells form the spongious zone,
gelatinous substance, proper nucleus and thoracic nucleus
(Clarc-Steiling column).
Lateral columns
in thoracic-lumbar portion contain autonomic cells that form lateral intermediate nucleus. In
intermediate central part medial
intermediate nucleus carry intermediate cells.
White substance
contains only neuron long processes (axons), the nerve fibbers which form
ascending (sensory, afferent) and descending (motor, efferent) pathways. White
substance divided by sulcuses and is arranged in three funiculi: anterior,
lateral, and posterior. Both anterior funiculi are communicated each other
by white commisura.
Fasciculi in the Posterior Funiculus comprises the fasciculus gracilis [tract of Goll] (lies next the posterior
median septum) and the fasciculus
cuneatus [tract of Burdach]
(laterally). They conduct impulses
of conscious muscle sense.
Pathways in the Anterior Funiculus (descending):
anterior corticospinal [pyramidal] tract, tectospinal tract, reticulospinal
tract, olivospinal tract, and vestibulospinal tract. Ascending
pathway - anterior spinothalamic tract.

Diagram of the principal fasciculi of the
spinal cord.
Ascending pathways in the Lateral Funiculus:
·
dorsal spinocerebellar tract (tract of Flechsig)
·
ventral spinocerebellar
tract (tract of Gowers)
·
lateral spinothalamic
·
tract
Medially they can find descending
pathways:
·
corticospinal [pyramidal] tract
·
rubrospinal tract (of Monakow)

Portion
of the spinal cord that carries two pairs of ventral and dorsal roots or one pair
of spinal nerves called segment.
There are 8 cervical, 12 thoracic, 5 lumbar, 5 sacral and 1 coccygeal segments.
Roots which exit from the spinal cord lower then second lumbar segment form
together with the filum termanale cauda
equina.
Gray
Substance (substantia grisea centralis).—The gray
substance consists of two symmetrical portions, one in each half of the medulla
spinalis: these are joined across the middle line by a transverse commissure of
gray substance, through which runs a minute canal, the central canal,
just visible to the naked eye. In a transverse section each half of the gray
substance is shaped like a comma or crescent, the concavity of which is
directed laterally; and these, together with the intervening gray commissure,
present the appearance of the letter H. An imaginary coronal plane through the
central canal serves to divide each crescent into an anterior or ventral,
and a posterior or dorsal column.
The
Anterior Column (columna anterior; anterior cornu), directed
forward, is broad and of a rounded or quadrangular shape. Its posterior part is
termed the base, and its anterior part the head, but these are not
differentiated from each other by any well-defined constriction. It is
separated from the surface of the medulla spinalis by a layer of white
substance which is traversed by the bundles of the anterior nerve roots. In the
thoracic region, the postero-lateral part of the anterior column projects
lateralward as a triangular field, which is named the lateral column (columna
lateralis; lateral cornu).
The
Posterior Column (columna posterior; posterior cornu) is long and
slender, and is directed backward and lateralward: it reaches almost as far as
the posterolateral sulcus, from which it is separated by a thin layer of white
substance, the tract of Lissauer. It consists of a base, directly
continuous with the base of the anterior horn, and a neck or slightly
constricted portion, which is succeeded by an oval or fusiform area, termed the
head, of which the apex approaches the posterolateral sulcus. The
apex is capped by a V-shaped or crescentic mass of translucent, gelatinous
neuroglia, termed the substantia gelatinosa of Rolando, which contains
both neuroglia cells, and small nerve cells. Between the anterior and posterior
columns the gray substance extends as a series of processes into the lateral
funiculus, to form a net-work called the formatio reticularis.
The
quantity of gray substance, as well as the form which it presents on transverse
section, varies markedly at different levels. In the thoracic region it is
small, not only in amount but relatively to the surrounding white substance. In
the cervical and lumbar enlargements it is greatly increased: in the latter,
and especially in the conus medullaris, its proportion to the white substance
is greatest (665).
In the cervical region its posterior column is comparatively narrow, while its
anterior is broad and expanded; in the thoracic region, both columns are
attenuated, and the lateral column is evident; in the lumbar enlargement, both
are expanded; while in the conus medullaris the gray substance assumes the form
of two oval masses, one in each half of the cord, connected together by a broad
gray commissure.
The
Throughout
the cervical and thoracic regions the central canal is situated in the anterior
third of the medulla spinalis; in the lumbar enlargement it is near the middle,
and in the conus medullaris it approaches the posterior surface. It is filled
with cerebrospinal fluid, and lined by ciliated, columnar epithelium, outside
of which is an encircling band of gelatinous substance, the substantia
gelatinosa centralis. This gelatinous substance consists mainly of
neuroglia, but contains a few nerve cells and fibers; it is traversed by
processes from the deep ends of the columnar ciliated cells which line the
central canal (667).
Structure
of the Gray Substance.—The gray substance consists of
numerous nerve cells and nerve fibers held together by neuroglia. Throughout
the greater part of the gray substance the neuroglia presents the appearance of
a sponge-like network, but around the central canal and on the apices of the
posterior columns it consists of the gelatinous substance already referred to.
The nerve cells are multipolar, and vary greatly in size and shape. They
consist of (1) motor cells of large size, which are situated in the anterior
horn, and are especially numerous in the cervical and lumbar enlargements; the
axons of most of these cells pass out to form the anterior nerve roots, but
before leaving the white substance they frequently give off collaterals, which
reënter and ramify in the gray substance. 113
(2) Cells of small or medium size, whose axons pass into the white matter,
where some pursue an ascending, and others a descending course, but most of
them divide in a T-shape manner into descending and ascending processes. They
give off collaterals which enter and ramify in the gray substance, and the
terminations of the axons behave in a similar manner. The lengths of these
axons vary greatly: some are short and pass only between adjoining spinal
segments, while others are longer and connect more distant segments. These
cells and their processes constitute a series of association or intersegmental
neurons (668),
which link together the different parts of the medulla spinalis. The axons of
most of these cells are confined to that side of the medulla spinalis in which
the nerve cells are situated, but some cross to the opposite side through the
anterior commissure, and are termed crossed commissural fibers. Some of
these latter end directly in the gray substance, while others enter the white
substance, and ascend or descend in it for varying distances, before finally
terminating in the gray substance. (3) Cells of the type II of Golgi, limited
for the most part to the posterior column, are found
also in the substantia gelatinosa of Rolando; their axons are short and
entirely confined to the gray substance, in which they break up into numerous
fine filaments. Most of the nerve cells are arranged in longitudinal columns,
and appear as groups on transverse section (Figs. 669, 670, 671).

Transverse sections of
the medulla spinalis at different
Nerve
Cells in the Anterior Column.—The nerve cells in the anterior
column are arranged in columns of varying length. The longest occupies the
medial part of the anterior column, and is named the antero-medial column:
it is well marked in C4, C5, again from C8 to L4, it disappears in L5 and S1
but is well marked in S2, S3 and S4 (Bruce). 114
Behind it is a dorso-medial column of small cells, which is not
represented in L5, S1, S2 nor below S4. Its axons probably pass into the dorsal
rami of the spinal nerves to supply the dorsal musculature of the spinal
column. In the cervical and lumbar enlargements, where the anterior column is
expanded in a lateral direction, the following additional columns are present,
viz.: (a) antero-lateral, which consists of two groups, one in
C4, C5, C6 the other in C6, C7, C8 in the cervical enlargement and of a group
from L2 to S2 in the lumbo-sacral enlargement; (b) postero-lateral,
in the lower five cervical, lower four lumbar, and upper three sacral segments;
(c) post-postero-lateral, in the last cervical, first thoracic,
and upper three sacral segments; and (d) a central, in the lower
four lumbar and upper two sacral segments. These cell groups are evidently
related to the nerve roots of the brachial and sacral plexuses and supply
fibers to the muscles of the arm and leg. Throughout the base of the anterior
column are scattered solitary cells, the axons of some of which form crossed
commissural fibers, while others constitute the motor fibers of the posterior
nerve roots. (See footnote, page 755.)
Nerve
Cells in the Lateral Column.—These form a column which is best
marked where the lateral gray column is differentiated, viz., in the thoracic
region; 115
but it can be traced throughout the entire length of the medulla spinalis in
the form of groups of small cells which are situated in the anterior part of
the formatio reticularis. In the upper part of the cervical region and lower
part of the medulla oblongata as well as in the third and fourth sacral
segments this column is again differentiated. In the medulla it is known as the
lateral nucleus. The cells of this column are fusiform or star-shaped,
and of a medium size: the axons of some of them pass into the
anterior nerve roots, by which they are carried to the sympathetic nerves: they
constitute the white rami and are sympathetic or visceral efferent fibers; they
are also known as preganglionic fibers of the sympathetic system; the
axons of others pass into the anterior and lateral funiculi, where they become
longitudinal.
Nerve
Cells in the Posterior Column.—1. The dorsal nucleus (nucleus
dorsalis; column of Clarke) occupies the medial part of the base of the
posterior column, and appears on the transverse section as a well-defined oval
area. It begins below at the level of the second or third lumbar nerve, and
reaches its maximum size opposite the twelfth thoracic nerve. Above the level
of the ninth thoracic nerve its size diminishes, and the column ends opposite
the last cervical or first thoracic nerve. It is represented, however, in the
other regions by scattered cells, which become aggregated to form a cervical
nucleus opposite the third cervical nerve, and a sacral nucleus in
the middle and lower part of the sacral region. Its cells are of medium size,
and of an oval or pyriform shape; their axons pass into the peripheral part of
the lateral funiculus of the same side, and there ascend, probably in dorsal
spinocerebellar (direct cerebellar) fasciculus. 2. The nerve
cells in the substantia gelatinosa of Rolando are arranged in three zones:
a posterior or marginal, of large angular or fusiform cells; an intermediate,
of small fusiform cells; and an anterior, of star-shaped cells. The axons of
these cells pass into the lateral and posterior funiculi, and there assume a
vertical course. In the anterior zone some Golgi cells are found whose short
axons ramify in the gray substance. 3. Solitary cells of varying form
and size are scattered throughout the posterior column. Some of these are
grouped to form the posterior basal column in the base of the posterior
column, lateral to the dorsal nucleus; the posterior basal column is
well-marked in the gorilla (Waldeyer), but is ill-defined in man. The axons of
its cells pass partly to the posterior and lateral funiculi of the same side,
and partly through the anterior white commissure to the lateral funiculus of
the opposite side. Golgi cells, type II, located in this region send axons to
the lateral and ventral columns.
A
few star-shaped or fusiform nerve cells of varying size are found in the
substantia gelatinosa centralis. Their axons pass into the lateral funiculus of
the same, or of the opposite side.
The
nerve fibers in the gray substance form a dense interlacement of minute fibrils
among the nerve cells. This interlacement is formed partly of axons which pass
from the cells in the gray substance to enter the white funiculi or nerve
roots; partly of the axons of Golgi’s cells which ramify only in the gray
substance; and partly of collaterals from the nerve fibers in the white
funiculi which, as already stated, enter the gray substance and ramify within
it.
White
Substance (substantia alba).—The white
substance of the medulla spinalis consists of medullated nerve fibers imbedded
in a spongelike net-work of neuroglia, and is arranged in three funiculi:
anterior, lateral, and posterior. The anterior funiculus lies between the
anterior median fissure and the most lateral of the anterior nerve roots: the
lateral funiculus between these nerve roots and the postero-lateral sulcus; and
the posterior funiculus between the postero-lateral and the posterior median
sulci (672).
The fibers vary greatly in thickness, the smallest being found in the
fasciculus gracilis, the tract of Lissauer, and inner part of the
lateral funiculus; while the largest are situated in the anterior funiculus,
and in the peripheral part of the lateral funiculus. Some of the nerve fibers
assume a more or less transverse direction, as for example those which cross
from side to side in the anterior white commissure, but the majority pursue a
longitudinal course and are divisible into (1) those connecting the medulla
spinalis with the brain and conveying impulses to or from the latter, and (2)
those which are confined to the medulla spinalis and link together its
different segments, i. e., intersegmental or association fibers.
Nerve
Fasciculi.—The longitudinal fibers are grouped into more or less
definite bundles or fasciculi. These are not recognizable from each other in
the normal state, and their existence has been determined by the following
methods: (1) A. Waller discovered that if a bundle of nerve fibers be cut, the
portions of the fibers which are separated from their cells rapidly degenerate
and become atrophied, while the cells and the parts of the fibers connected with
them undergo little alteration. 116
This is known as Wallerian degeneration. Similarly, if a group of nerve
cells be destroyed, the fibers arising from them undergo degeneration. Thus, if
the motor cells of the cerebral cortex be destroyed, or if the fibers arising
from these cells be severed, a descending degeneration from the seat of
injury takes place in the fibers. In the same manner, if a spinal ganglion be
destroyed, or the fibers which pass from it into the medulla spinalis be cut,
an ascending degeneration will extend along these fibers. (2)
Pathological changes, especially in man, have given important information by
causing ascending and descending degenerations. (3) By tracing the development
of the nervous system, it has been observed that at first the nerve fibers are
merely naked axis-cylinders, and that they do not all acquire their medullary
sheaths at the same time; hence the fibers can be grouped into different
bundles according to the dates at which they receive their medullary sheaths.
(4) Various methods of staining nervous tissue are of great value in tracing
the course and mode of termination of the axis-cylinder processes.
Fasciculi
in the Anterior Funiculus.—Descending Fasciculi.—The
anterior cerebrospinal (fasciculus cerebrospinalis anterior; direct
pyramidal tract), which is usually small, but varies inversely in size with
the lateral cerebrospinal fasciculus. It lies close to the anterior median
fissure, and is present only in the upper part of the medulla spinalis;
gradually diminishing in size as it descends, it ends about the middle of the
thoracic region. It consists of descending fibers which arise from
cells in the motor area of the cerebral hemisphere of the same side, and which,
as they run downward in the medulla spinalis, cross in succession through the
anterior white commissure to the opposite side, where they end, either directly
or indirectly, by arborizing around the motor cells in the anterior column. A
few of its fibers are said to pass to the lateral column of the same side and
to the gray matter at the base of the posterior column. They conduct voluntary
motor impulses from the precentral gyrus to the motor centers of the cord.
The
vestibulospinal fasciculus, situated chiefly in the marginal part of the
funiculus and mainly derived from the cells of Deiters’ nucleus, of the same
and the opposite side, i. e., the chief terminal nucleus of the
vestibular nerve. Fibers are also contributed to this fasciculus from scattered
cells of the articular formation of the medulla oblongata, the pons and the
mid-brain (tegmentum). The other terminal nuclei of the vestibular nerve also
contribute fibers. In the brain stem these fibers form part of the median
longitudinal bundle. The fasciculus can be traced to the sacral region. Its
terminals and collaterals end either directly or indirectly among the motor
cells of the anterior column. This fasciculus is probably concerned with
equilibratory reflexes.
The
tectospinal fasciculus, situated partly in the anterior and partly in
the lateral funiculus, is mainly derived from the opposite superior colliculus
of the mid-brain. The fibers from the superior colliculus cross the median
raphé in the fountain decussation of Meynert and descend as the ventral
longitudinal bundle in the reticular formation of the brain-stem. Its
collaterals and terminals end either directly or indirectly among the motor
cells of the anterior column of the same side. Since the superior colliculus is
an important visual reflex center, the tectospinal fasciculus is probably
concerned with visual reflexes.
Ascending
Fasciculi.—The ventral spinothalamic fasciculus, situated in the
marginal part of the funiculus and intermingled more or less with the
vestibulo-spinal fasciculus, is derived from cells in the posterior column or
intermediate gray matter of the opposite side. Their axons cross in the
anterior commissure. This is a somewhat doubtful fasciculus and its fibers are
supposed to end in the thalamus and to conduct certain of the touch impulses.
The
remaining fibers of the anterior funiculus constitute what is termed the anterior
proper fasciculus (fasciculus anterior proprius; anterior basis bundle).
It consists of (a) longitudinal intersegmental fibers which arise from
cells in the gray substance, more especially from those of the medial group of
the anterior column, and, after a longer or shorter course, reënter the
gray substance; (b) fibers which cross in the anterior white commissure
from the gray substance of the opposite side.
Fasciculi
in the Lateral Funiculus.—1. Descending Fasciculi.—(a)
The lateral cerebrospinal fasciculus (fasciculus cerebrospinalis
lateralis; crossed pyramidal tract) extends throughout the entire length of
the medulla spinalis, and on transverse section appears as an oval area in
front of the posterior column and medial to the cerebellospinal. Its fibers
arise from cells in the motor area of the cerebral hemisphere of the opposite
side. They pass downward in company with those of the anterior cerebrospinal
fasciculus through the same side of the brain as that from which they
originate, but they cross to the opposite side in the medulla oblongata and
descend in the lateral funiculus of the medulla spinalis.
It
is probable 117
that the fibers of the anterior and lateral cerebrospinal fasciculi are not
related in this direct manner with the cells of the anterior column, but
terminate by arborizing around the cells at the base of the posterior column
and the cells of Clarke’s column, which in turn link them to the motor cells in
the anterior column, usually of several segments of the cord. In consequence of
these interposed neurons the fibers of the cerebrospinal
fasciculi correspond not to individual muscles, but to associated groups of
muscles.
The
anterior and lateral cerebrospinal fasciculi constitute the motor fasciculi of
the medulla spinalis and have their origins in the motor cells of the cerebral
cortex. They descend through the internal capsule of the cerebrum, traverse the
cerebral peduncles and pons and enter the pyramid of the medulla oblongata. In
the lower part of the latter about two-thirds of them cross the middle line and
run downward in the lateral funiculus as the lateral cerebrospinal fasciculus,
while the remaining fibers do not cross the middle line, but are continued into
the same side of the medulla spinalis, where they form the anterior
cerebrospinal fasciculus. The fibers of the latter, however, cross the middle
line in the anterior white commissure, and thus all the motor fibers from one
side of the brain ultimately reach the opposite side of the medulla spinalis.
The proportion of fibers which cross in the medulla oblongata is not a constant
one, and thus the anterior and lateral cerebrospinal fasciculi vary inversely
in size. Sometimes the former is absent, and in such cases it may be presumed
that the decussation of the motor fibers in the medulla oblongata has been
complete. The fibers of these two fasciculi do not acquire their medullary
sheaths until after birth. In some animals the motor fibers are situated in the
posterior funiculus.
(b)
The rubrospinal fasciculus (Monakow) (prepyramidal tract), lies
on the ventral aspect of the lateral cerebrospinal fasciculus and on transverse
section appears as a somewhat triangular area. Its fibers descend from the
mid-brain, where they have their origin in the red nucleus of the tegmentum of
the opposite side. Its terminals and collaterals end either directly or
indirectly in relation with the motor cells of the anterior column. The
rubrospinal fasciculus is supposed to be concerned with cerebellar reflexes
since fibers which pass from the cerebellum through the superior peduncle send
many collaterals and terminals to the red nucleus.
(c)
The olivospinal fasciculus (Helweg) arises in the vicinity of the
inferior olivary nucleus in the medulla oblongata, and is seen only in the
cervical region of the medulla spinalis, where it forms a small triangular area
at the periphery, close to the most lateral of the anterior nerve roots. Its
exact origin and its mode of ending have not yet been definitely made out.
2.
Ascending Fasciculi.—(a) The dorsal spinocerebellar fasciculus (fasciculus
cerebellospinalis; direct cerebellar tract of Flechsig) is situated at the
periphery of the posterior part of the lateral funiculus, and on transverse
section appears as a flattened band reaching as far forward as a line drawn
transversely through the central canal. Medially, it is in contact with the
lateral cerebrospinal fasciculus, behind, with the fasciculus of Lissauer. It
begins about the level of the second or third lumbar nerve, and increasing in
size as it ascends, passes to the vermis of the cerebellum through the inferior
peduncle. Its fibers are generally regarded as being formed by the axons of the
cells of the dorsal nucleus (Clarke’s column); they receive their medullary
sheaths about the sixth or seventh month of fetal life. Its fibers are supposed
to conduct impulses of unconscious muscle sense.
The
superficial antero-lateral fasciculus (tract of Gowers) consists
of four fasciculi, the ventral spinocerebellar, the lateral spinothalamic, the
spinotectal and the ventral spinothalamic.
(b)
The ventral spinocerebellar fasciculus (Gowers) skirts the
periphery of the lateral funiculus in front of the dorsal spinocerebellar
fasciculus. In transverse section it is shaped somewhat like a comma, the
expanded end of which lies in front of the dorsal spinocerebellar fasciculus
while the tail reaches forward into the anterior funiculus. Its fibers come
from the same but mostly from the opposite side of the medulla spinalis and
cross both in the anterior white commissure and in the gray commissure; they
are probably derived from the cells of the dorsal nucleus and from other cells
of the posterior column and the intermediate portion of
the gray matter. The ventral spinocerebellar fasciculus begins about the level
of the third pair of lumbar nerves, and can be followed into the medulla
oblongata and pons almost to the level of the inferior colliculus where it
crosses over the superior peduncle and then passes backward along its medial
border to reach the vermis of the cerebellum. In the pons it lies along the
lateral edge of the lateral lemniscus. Some of its fibers join the dorsal
spinocerebellar fasciculus at the level of the inferior peduncle and pass with
them into the cerebellum. Other fibers are said to continue upward in the
dorso-lateral part of the tegmentum of the mid-brain probably as far as the
thalamus.
(c)
The lateral spinothalamic fasciculus is supposed to come from cells in
the dorsal column and the intermediate gray matter whose axons cross in the
anterior commissure to the opposite lateral funiculus where they pass upward on
the medial side of the ventral spinocerebellar fasciculus; on reaching the
medulla oblongata they continue in the formatio reticularis near the median
fillet and probably terminate in the ventro-lateral region of the thalamus. It
is supposed to conduct impulses of pain and temperature. The lateral and
ventral spinothalamic fasciculi are sometimes termed the secondary sensory
fasciculus or spinal lemniscus.
(d)
The spinotectal fasciculus is supposed to arise in the dorsal column and
terminate in the (inferior ?) and superior colliculi. It is situated ventral to
the lateral spinothalamic fasciculus, but its fibers are more or less intermingled
with it. It is also known as the spino-quadrigeminal system of Mott. In
the brain-stem the fibers run lateral from the inferior olive, ventro-lateral
from the superior olive, then ventro-medial from the spinal tract of the
trigeminal; the fibers come to lie in the medial portion of the lateral
lemniscus.
(e)
The fasciculus of Lissauer is a small strand situated in relation to the
tip of the posterior column close to the entrance of the posterior nerve roots.
It consists of fine fibers which do not receive their medullary sheaths until
toward the close of fetal life. It is usually regarded as being formed by some
of the fibers of the posterior nerve roots, which ascend for a short distance
in the tract and then enter the posterior column, but since its fibers are
myelinated later than those of the posterior nerve roots, and do not undergo
degeneration in locomotor ataxia, they are probably intersegmental in
character.
In
addition the fasciculus or tract of Lissauer contains great numbers of
fine non-medullated fibers derived mostly from the dorsal roots but partly
endogenous in origin. These fibers are intimately related to the substantia
gelatinosa which is probably the terminal nucleus. The non-medullated fibers
ascend or descend for short distances not exceeding one or two segments, but
most of them enter the substantia gelatinosa at or near the level of their
origin. Ransom 118
suggests that these non-medullated fibers and the substantia gelatinosa are
concerned with the reflexes associated with pain impulses.
(f)
The lateral proper fasciculus (fasciculus lateralis proprius; lateral
basis bundle) constitutes the remainder of the lateral column, and is
continuous in front with the anterior proper fasciculus. It consists chiefly of
intersegmental fibers which arise from cells in the gray substance, and, after
a longer or shorter course, reënter the gray substance and ramify in it.
Some of its fibers are, however, continued upward into the brain under the name
of the medial longitudinal fasciculus.
Fasciculi
in the Posterior Funiculus.—This funiculus comprises two main
fasciculi, viz., the fasciculus gracilis, and the fasciculus
cuneatus. These are separated from each other in the cervical and upper
thoracic regions by the postero-intermediate septum, and consist mainly of
ascending fibers derived from the posterior nerve roots.
The
fasciculus gracilis (tract of Goll) is wedge-shaped on transverse
section, and lies next the posterior median septum, its base being at the
surface of the medulla spinalis, and its apex directed
toward the posterior gray commissure. It increases in size from below upward,
and consists of long thin fibers which are derived from the posterior nerve
roots, and ascend as far as the medulla oblongata, where they end in the
nucleus gracilis.
The
fasciculus cuneatus (tract of Burdach) is triangular on
transverse section, and lies between the fasciculus gracilis and the posterior
column, its base corresponding with the surface of the medulla spinalis. Its
fibers, larger than those of the fasciculus gracilis, are mostly derived from
the same source, viz., the posterior nerve roots. Some ascend for only a short
distance in the tract, and, entering the gray matter, come into close
relationship with the cells of the dorsal nucleus; while others can be traced
as far as the medulla oblongata, where they end in the gracile and cuneate
nuclei.
The
fasciculus gracilis and fasciculus cuneatus conduct (1) impulses of conscious
muscle sense, neurons of the second order from the nucleus gracilis and nucleus
cuneatus, pass in the median lemniscus to the thalamus and neurons of the third
order from the thalamus to the cerebral cortex; (2)
impulses of unconscious muscle sense, via neurons of the second order
from the nucleus gracilis and nucleus cuneatus pass in the internal and
external arcuate fibers of the medulla oblongata to the inferior peduncle and
through it to the cerebellum; (3) impulses of tactile discrimination, via
neurons of the second order from the nucleus cuneatus and nucleus gracilis pass
in the median lemniscus to the thalamus, neurons of the third order pass from
the thalamus to the cortex.
The
Posterior Proper Fasciculus (posterior ground bundle; posterior basis
bundle) arises from cells in the posterior column; their axons bifurcate
into ascending and descending branches which occupy the ventral part of the
funiculus close to the gray column. They are intersegmental and run for varying
distances sending off collaterals and terminals to the gray matter.
Some
descending fibers occupy different parts at different levels. In the cervical
and upper thoracic regions they appear as a comma-shaped fasciculus in
the lateral part of the fasciculus cuneatus, the blunt end of the comma being
directed toward the posterior gray commissure; in the lower thoracic region
they form a dorsal peripheral band on the posterior surface of the
funiculus; in the lumbar region, they are situated by the side of the posterior
median septum, and appear on section as a semi-elliptical bundle, which,
together with the corresponding bundle of the opposite side, forms the oval
area of Flechsig; while in the conus medullaris they assume the form of a triangular
strand in the postero-medial part of the fasciculus gracilis. These
descending fibers are mainly intersegmental in character and derived from cells
in the posterior column, but some consist of the descending branches of the
posterior nerve roots. The comma-shaped fasciculus was supposed to belong to
the second category, but against this view is the fact that it does not undergo
descending degeneration when the posterior nerve roots are destroyed.
Roots
of the Spinal Nerves.—As already stated, each spinal nerve
possesses two roots, an anterior and a posterior, which are
attached to the surface of the medulla spinalis opposite the corresponding
column of gray substance (675);
their fibers become medullated about the fifth month of fetal life.
The
Anterior Nerve Root (radix anterior) consists of efferent fibers,
which are the axons of the nerve cells in the ventral part of the anterior and
lateral columns. A short distance from their origins, these axons are invested
by medullary sheaths and, passing forward, emerge in two or three irregular
rows over an area which measures about
The
Posterior Root (radix posterior) comprises some six or eight
fasciculi, attached in linear series along the postero-lateral sulcus. It
consists of afferent fibers which arise from the nerve cells in a spinal
ganglion. Each ganglion cell gives off a single fiber which divides in a
T-shaped manner into two processes, medial and lateral. The lateral processes
extend to the sensory end-organs of the skin, muscles, tendons, joints, etc. (somatic
receptors), and to the sensory end-organs of the viscera (visceral receptors).
The medial processes of the ganglion cells grow into the medulla spinalis as the
posterior roots of the spinal nerves.
The
posterior nerve root enters the medulla spinalis in three chief bundles,
medial, intermediate, and lateral. The medial strand passes directly
into the fasciculus cuneatus: it consists of coarse fibers, which acquire their
medullary sheaths about the fifth month of intrauterine life; the intermediate
strand consists of coarse fibers, which enter the gelatinous substance of
Rolando; the lateral is composed of fine fibers, which assume a
longitudinal direction in the tract of Lissauer, and do not acquire their
medullary sheaths until after birth. In addition to these medullated fibers
there are great numbers of non-medullated fibers which enter with the lateral
bundle. They are more numerous than the myelinated fibers. They arise from the
small cells of the spinal ganglia by T-shaped axons similar to the myelinated.
They are distributed with the peripheral nerves chiefly to the skin, only a few
are found in the nerves to the muscles. 119
Having
entered the medulla spinalis, all the fibers of the posterior nerve roots
divide into ascending and descending branches, and these in their turn give off
collaterals which enter the gray substance (676).
The descending fibers are short, and soon enter the gray substance. The
ascending fibers are grouped into long, short, and intermediate: the long
fibers ascend in the fasciculus cuneatus and fasciculus gracilis as far as the
medulla oblongata, where they end by arborizing around the cells of the cuneate
and gracile nuclei; the short fibers run upward for a distance of only 5 or
The
long fibers of the posterior nerve roots pursue an oblique course upward, being
situated at first in the lateral part of the fasciculus cuneatus: higher up,
they occupy the middle of this fasciculus, having been displaced by the
accession of other entering fibers; while still higher, they ascend in the
fasciculus gracilis. The upper cervical fibers do not reach this fasciculus,
but are entirely confined to the fasciculus cuneatus. The localization of these
fibers is very precise: the sacral nerves lie in the medial part of the
fasciculus gracilis and near its periphery, the lumbar nerves lateral to them,
the thoracic nerves still more laterally; while the cervical nerves are confined
to the fasciculus cuneatus.
Prepared by
Galytska-Harhalis
O.Ya.